|
||||
6. Как работает кора головного мозгаПытаться разгадать, как работает кора головного мозга, все равно, что собирать пазлы – головоломку, в которой картинку складывают из множества отдельных кусочков. Пазлы можно собирать двумя способами. Первый – от целого к частному – предполагает, что у вас есть образец картинки, которая должна получиться. Начав собирать с какой-либо части изображения, вы подыскиваете подходящие кусочки и шаг за шагом восстанавливаете его целиком. Второй подход – от частного к целому – подразумевает сосредоточенность на отдельных кусочках. Вы ищете в отдельных фрагментах картины общие детали и пытаетесь соотнести их друг с другом. При отсутствии подсказки в виде готового изображения единственно возможным способом собирания пазлов является второй. Головоломка под условным названием «Как понять мозг» особенно трудна. Поскольку единого подхода к пониманию мозга долгое время вообще не существовало, исследователи были вынуждены прибегать к методу от частного к целому. С таким уровнем сложности, как мозг человека, данная задача была по силам лишь Геркулесу. Только представьте себе: каждый из многих тысяч фрагментов – двухсторонний, причем не существует однозначных его интерпретаций, и только одна из сторон каждого кусочка имеет отношение к головоломке. Края всех кусочков неровные (словно они были неаккуратно вырезаны), поэтому вы не можете судить с полной уверенностью, правильно ли соединяете каждую пару. Многие из имеющихся у вас фрагментов вообще не нужны для составления этого гигантского изображения, их наличие в общей куче лишь усложняет вам задачу. Кроме того, по почте вам постоянно присылают все новые и новые кусочки, причем некоторые из них предназначены для того, чтобы заменить старые. Автор головоломки как бы говорит вам: «Я знаю, что вы несколько лет бились над тем, как соединить все эти фрагменты, но оказалось, что они не имеют отношения к моей картинке. Вы уж извините. Вот вам новые, до следующего раза». И самое главное: вы даже не представляете себе, каким должен быть конечный результат. Или того хуже – у вас есть определенные соображения, но они ошибочны. Аналогия с головоломкой-пазлами прекрасно иллюстрирует трудности, с которыми мы сталкиваемся на пути создания единой теории неокортекса и интеллекта. Кусочки головоломки – это биологические и поведенческие данные, накопленные учеными за сто с лишним лет. Результаты новых исследований добавляют в головоломку новые кусочки. Иногда сведения, предложенные разными учеными, крайне противоречивы. А поскольку любые данные можно истолковывать по-разному, то единого мнения не существует практически ни по какому вопросу. Ввиду невозможности подхода от целого к частному цель научных изысканий зачастую остается невыясненной, и, что не менее важно, ученые порой не имеют представления о том, как интерпретировать массивы накопленной информации. Изучая мозг с помощью подхода от частного к целому, мы попросту зашли в тупик. Очевидно, необходимо очертить рамки для подхода от целого к частному. Такими рамками может послужить модель «Память-запоминание». Она подскажет нам, как сложить вместе все кусочки головоломки в целую картинку. Для осуществления прогностической функции неокортексу нужна определенная технология запоминания и сохранения информации о последовательностях событий. Для прогнозирования вероятных событий кора головного мозга должна использовать (а значит, формировать) инвариантные представления. Ваш мозг должен создавать и сохранять модель реального мира так, чтобы при восприятии последнего в разных обстоятельствах изменения компенсировались. Разобравшись в том, чем должна заниматься кора головного мозга, мы сможем понять ее строение, особенно шестислойную иерархическую структуру. В данной книге такое видение рамок исследования интеллекта будет представлено впервые. Иногда я буду забираться в научные дебри, что, возможно, некоторым читателям покажется слишком сложным. Многие из концепций будут незнакомы даже специалистам-нейрологам. И все же, приложив определенные усилия, любой читатель, как минимум, сможет понять основы нового подхода. (Главы 7 и 8 гораздо меньше насыщены техническими подробностями – в них речь пойдет о более широком применении предложенной теории.) Прежде всего мы рассмотрим биологические предпосылки, на которых основана запоминающе-прогностическая гипотеза. Это поможет нам отбросить многие ненужные кусочки воображаемых пазлов, тем более что нам известно: лишь немногие из элементов действительно необходимы для воссоздания полной картины. Как только мы поставим перед собой четкую цель и разберемся, что же нужно искать, задача станет разрешимой. Хочу отметить, что предлагаемые мной рамки исследования далеко не окончательны. У меня до сих пор нет ответов на многие вопросы. Но, основываясь на дедуктивном методе поиска решений, результатах многочисленных экспериментов, проведенных мною во многих лабораториях, а также на знаниях анатомии, мне удалось обнаружить интересные факты, которыми я намерен поделиться с вами. На протяжении последнего десятилетия ученые-нейробиологи занимались исследованием идей, подобных моим. Обычно они использовали другую терминологию и, насколько мне известно, не пытались создать на основе полученных разрозненных сведений общую теорию. В отчетах об исследованиях они также упоминают обработку данных по принципу от целого к частному и от частного к целому, говорят о том, как нервные сигналы передаются между сенсорными областями мозга, и отмечают важную роль инвариантных репрезентаций. Так, Габриэль Крейман и Кристофер Кох, нейробиологи, работающие в Калифорнийском технологическом институте, и Айзек Фрейд из Калифорнийского университета в Лос-Анджелесе открыли нервные клетки, посылающие разряд в том случае, когда человек видит лицо Билла Клинтона. Одна из моих целей – объяснить вам, откуда появились эти нейроны Билла Клинтона. Естественно, создание каждой теории сопряжено с формулированием гипотез, которые следует проверять в лабораторных условиях. (В приложении к данной книге я перечислю несколько таких гипотез.) Теперь, когда у нас есть цель и мы знаем, что ищем, невероятно сложная, на первый взгляд, система уже не будет казаться нам такой уж замысловатой. Ниже в этой главе мы детально углубимся в модель «Память-предсказание», характеризующую работу неокортекса. Вначале рассмотрим структуру и наиболее общие функции коры головного мозга, а затем двинемся дальше – обратимся к частным вопросам и проанализируем, как они вписываются в целостную картину. 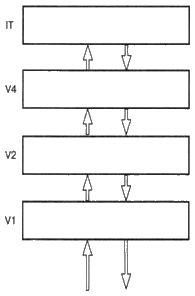 Рис. 6.1. Первые четыре зоны в распознавании объектов Инвариантные представленияПомните ли вы, как я назвал кору головного мозга тканью из нервных клеток размером со столовую салфетку и толщиной в шесть визитных карточек, в которой связи между разными зонами создают определенную иерархическую структуру? Сейчас я хочу проиллюстрировать иерархическую структуру неокортекса иначе. Представьте, что мы разрезали эту салфетку на несколько частей, соответствующих зонам, специализирующимся на выполнении определенных задач, и наложили полученные кусочки друг на друга. У нас вышел своеобразный слоеный, или «блинный» пирог. Разрезав «пирог», получим срез (см. рис. 6.3). Разумеется, в действительности неокортекс выглядит по-другому, но благодаря этой картинке вы составите четкое представление о передвижении потоков информации. На рис. 6.1 показано, как информационные сигналы поступают в низшую зону коры головного мозга и передвигаются вверх от зоны к зоне. Обратите внимание, что информационные потоки циркулируют в обоих направлениях. Рис. 6.1 представляет первые четыре зрительные зоны, вовлеченные в распознание объектов окружающего мира. С их помощью вы узнаете кошку, церковь, свою мать, Большую китайскую стену и так далее. Биологи обозначают эти зоны V1, V2, V4 и IT. Входной визуальный сигнал показан стрелкой под зоной V1. Зрительная информация с сетчатки ваших глаз передается к V1. Этот входной поток информации можно представить как непрерывно изменяющиеся последовательности сигналов, которые передаются приблизительно по миллиону аксонов, составляющих ваш зрительный нерв. Ранее мы уже упоминали о пространственно-временных сигналах, однако сейчас стоит освежить эти знания, поскольку мы будем часто к ним обращаться. Кора головного мозга состоит из функциональных зон, специализирующихся на отдельных задачах. Эти зоны связаны друг с другом большими пучками аксонов, или нервных волокон, передающих информацию из одной зоны в другую. Каждую секунду какой-то пучок волокон посылает электрический разряд, называемый потенциалом действия, или импульсом, в то время как другие пучки молчат. Общая активность пучка волокон называется паттерном (последовательностью сигналов). Паттерн, полученный зоной V1, будет пространственным, если ваши глаза на мгновение остановились на объекте, и временным, если они его разглядывают. Примерно трижды в секунду ваши глаза осуществляют скачкообразное движение – саккаду, после чего следует пауза – фиксация. Если бы вы участвовали в эксперименте и исследователь при помощи специального устройства отслеживал движения ваших глаз, вы бы удивились, узнав, насколько резки саккады, при том что воспринимаемое изображение кажется вам постоянным и устойчивым. На рис. 6.2 а показано, как двигались глаза одного из участников эксперимента, когда он рассматривал изображение лица. Обратите внимание, что фиксации вовсе не беспорядочны. А теперь представьте, что вы могли бы видеть паттерн, поступающий к зоне V1 этого человека. Он полностью меняется с каждой саккадой. Значит, несколько раз в секунду зрительная зона коры головного мозга получает совершенно новую последовательность сигналов.  Рис. 6.2. а) Траектория саккад при рассматривании лица человека Возможно, вы подумаете: «Ну ладно, ведь все равно это лицо остается одним и тем же, несмотря на сдвиги». Доля истины в этом есть, но намного меньшая, чем вы себе представляете. Световые рецепторы на сетчатке глаза распределены неравномерно. Их плотность, очень высокая в центральной ямке, постепенно уменьшается по направлению к периферии глаза. А нервные клетки коры головного мозга, напротив, распределены равномерно. В результате изображение, переданное с сетчатки в первичную зрительную зону V1, очень искажено. Если вы, рассматривая одно и то же лицо, направите взгляд на нос, а потом сконцентрируете его на глазе, зрительный входной сигнал будет очень отличаться, как будто вы смотрите на это лицо через немилосердно дергающийся «рыбий глаз»[15]. Тем не менее при взгляде на лицо оно не кажется вам искаженным; изображение не «прыгает» у вас перед глазами. Скорее всего, вы вообще не осознаете, что получаемые от сетчатки визуальные сигналы изменяются, более того – так сильно изменяются. Вы просто видите «лицо» (на рис. 6.2 б показан эффект взгляда на прибрежный пейзаж через линзу «рыбий глаз»). По сути, это переформулирование загадки об инвариантном представлении, о которой мы беседовали в главе 4.  Рис. 6.2. 6) искажение, вызванное неравномерным распределением рецепторов в сетчатке глаза Возможно, вы подумаете: «Ну ладно, ведь все равно это лицо остается одним и тем же, несмотря на сдвиги». Доля истины в этом есть, но намного меньшая, чем вы себе представляете. Световые рецепторы на сетчатке глаза распределены неравномерно. Их плотность, очень высокая в центральной ямке, постепенно уменьшается по направлению к периферии глаза. А нервные клетки коры головного мозга, напротив, распределены равномерно. В результате изображение, переданное с сетчатки в первичную зрительную зону V1, очень искажено. Если вы, рассматривая одно и то же лицо, направите взгляд на нос, а потом сконцентрируете его на глазе, зрительный входной сигнал будет очень отличаться, как будто вы смотрите на это лицо через немилосердно дергающийся «рыбий глаз»[16]. Тем не менее при взгляде на лицо оно не кажется вам искаженным; изображение не «прыгает» у вас перед глазами. Скорее всего, вы вообще не осознаете, что получаемые от сетчатки визуальные сигналы изменяются, более того – так сильно изменяются. Вы просто видите «лицо» (на рис. 6.2 б показан эффект взгляда на прибрежный пейзаж через линзу «рыбий глаз»). По сути, это переформулирование загадки об инвариантном представлении, о которой мы беседовали в главе 4. Вы воспринимаете не то, что видит зона V1. Откуда ваш мозг знает, что вы смотрите на одно и то же лицо, и почему вашему восприятию совершенно не мешает, что входные сигналы постоянно меняются, более того, они очень искажены? Поместив в зону V1 электрод и наблюдая за «поведением» отдельных клеток, мы обнаружим, что каждый нейрон посылает разряд только в ответ на зрительный сигнал из крошечной части сетчатки. Такой опыт по изучению зрительной функции проводили много раз. Каждый нейрон зоны V1 привязан к своему рецептивному полю, которое является очень ограниченным по сравнению с общим полем зрения. Нервные клетки зоны V1 ничего не знают о лицах, автомобилях, книгах или других объектах, постоянно встречающихся на вашем пути. Все, что они знают, – это крошечная, как укол булавки, часть всего мира перед вашими глазами. Каждая клетка зоны V1 настроена на восприятие определенных видов входных сигналов. Например, некий нейрон бурно реагирует, когда в его рецептивное поле попадает линия или угол в тридцать градусов. Сам по себе такой угол не имеет особенного значения. Он может быть частью какого угодно предмета – доской паркета, обломком дерева, частью буквы М. При каждой новой фиксации рецептивное поле клетки исследует новую часть видимого пространства. При разных фиксациях поток импульсов, посылаемый клеткой, может быть сильнее или слабее, а может и вовсе отсутствовать, т. е. при каждой саккаде активность клеток в зоне V1 изменяется. А вот если мы введем электрод в верхнюю зону IT (см. рис. 6.1), то обнаружим нечто совершенно невероятное. Мы увидим, что некоторые клетки указанной зоны возбуждаются и остаются активными, когда в поле зрения человека появляются целые объекты. Например, мы можем найти клетку, энергично реагирующую каждый раз, когда в поле зрения появляется лицо. Эта клетка будет активной до тех пор, пока лицо присутствует в любой точке поля зрения, под любым ракурсом и при любом освещении. Она не включается-выключается при каждой последующей саккаде, как это делают клетки зоны V1. Рецептивное поле такой клетки покрывает большую часть зрительного пространства, и она возбуждается всякий раз, когда человек видит лица. Остановимся на этом подробнее. Какие изменения характеризуют четыре уровня зон коры головного мозга? По мере продвижения от сетчатки до зоны IT изменчивые, пространственно специфические, ориентированные на распознание мелких деталей нейроны сменяются нейронами высших зон – пространственно неспецифическими, постоянно активными и способными распознавать целые объекты. Нейрон из зоны IT сообщает нам, что где-то в поле нашего зрения присутствует лицо. Эту нервную клетку называют клеткой лица, и она активизируется независимо от того, стоит ли хозяин лица прямо, склонил ли он голову или повернулся к вам в профиль. Это часть инвариантной репрезентации для объекта «лицо». Да, описать данную схему несложно. Проходим быстренько четыре стадии, и вот вам лицо. Но ни одна компьютерная программа, ни одна математическая формула не в состоянии обеспечить решение подобной задачи с той же надежностью и универсальностью, как это делает человеческий мозг. Нам известно, что мозг решает эту задачу за несколько шагов, значит, разгадка не должна быть слишком сложной. Одной из задач данной главы является поиск объяснения возникновения этой «клетки лица» (а также «клетки Билла Клинтона» и любой другой). Мы достигнем своей цели, но прежде нам следует обработать массу информации. Как говорится, всему свое время. Как видно из рис. 6.1, по сети обратных связей информация поступает из высших зон неокортекса в низшие. Эти обратные связи представлены пучками аксонов, которые проходят от высшей зоны IT к низшим зонам V4, V2, V1. Примечательно, что количество обратных связей в зрительной коре головного мозга превышает количество восходящих связей. Долгие годы ученые-нейробиологи игнорировали эти обратные связи, что вполне объяснимо: если вы ограничиваете свои задачи изучением того, как мозг принимает входную информацию, подвергает ее обработке, а потом на основе этого моделирует поведение, то в рассмотрении обратных связей, казалось бы, совсем нет необходимости. В таком случае фокус внимания исследователей прикован к прямым связям – проведению нервных импульсов от сенсорных зон головного мозга в моторные зоны. Однако если принять за основу идею, что ключевой функцией неокортекса является прогнозирование, то без изучения обратных связей не обойтись. Мозгу необходимо отсылать информацию из высших нервных центров обратно к тем зонам, от которых он ее получил: для того чтобы делать прогнозы, надо сравнивать реальные события и ожидания. То, что на самом деле произошло, идет снизу вверх, а то, чего вы ожидаете, спускается сверху вниз. Подобный круговой процесс происходит во всех сенсорных зонах коры головного мозга. На рис. 6.3 изображены аналогичные «слоеные пироги» для тактильного, слухового и зрительного восприятия. Также показаны высшие или ассоциативные зоны коры головного мозга, которые получают и анализируют информационные стимулы от нескольких органов чувств, в данном случае – слуха, зрения и осязания. 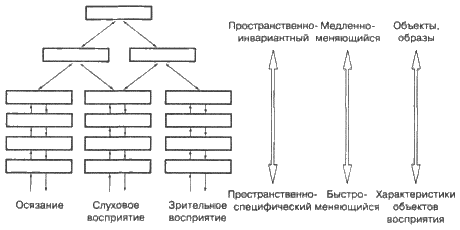 Рис. 6.3. Формирование инвариантных репрезентаций при осязании, слуховом и зрительном восприятии Если рис. 6.1 представляет общеизвестную взаимосвязь четырех хорошо изученных зон коры головного мозга, то рис. 6.3 – это условная диаграмма, целью которой не является отображение фактических зон неокортекса. В действительности десятки зон коры головного мозга человека взаимосвязаны друг с другом самыми различными способами. Большая часть неокортекса образована ассоциативными зонами. (Я использую схематические рисунки, чтобы помочь вам лучше разобраться в излагаемом материале. И надеюсь, что они не введут вас в заблуждение.) Переход от быстроменяющихся к медленноменяющимся и от пространственно-специфических к пространственно-инвариантным нейронам был доказан многочисленными результатами исследований зрительного восприятия. В отношении других сенсорных зон данных значительно меньше, но нейробиологи утверждают, что так же устроены и все остальные сенсорные зоны коры головного мозга. Рассмотрим, например, слуховое восприятие. Когда вы слышите устную речь, изменения звуковых колебаний происходят очень быстро. Так же быстро изменяются паттерны, поступающие в первичную слуховую зону А1. Но если бы мы могли поместить электрод в высшую зону неокортекса, ответственную за слуховое восприятие, то обнаружили бы инвариантные клетки, реагирующие на определенные слова и даже фразы. Вполне вероятно, что в вашей слуховой зоне коры головного мозга существует группа клеток, активизирующихся всякий раз, когда вы слышите слово «спасибо», или группа клеток, реагирующих на приветствие «доброе утро». Такие клетки остаются активными на протяжении всего времени, пока звучит фраза, при условии, конечно, что вы ее воспринимаете. Последовательности сигналов, попадающие в низшую слуховую зону, могут очень сильно отличаться друг от друга. Одно и то же слово можно произнести с разными акцентами, разным тоном и в разном темпе. Чем выше мы поднимаемся по иерархии слуховых зон коры головного мозга, тем менее важными становятся эти низкоуровневые различия. Слово идентифицируется согласно его значению, независимо от акустических эффектов. То же самое справедливо и в отношении музыки. Вы можете услышать одну и ту же песню, сыгранную на фортепиано, на кларнете или в исполнении ребенка. В каждом из перечисленных случаев ваша зона А1 получает совершенно различные звуковые сигналы. Тем не менее электрод, введенный в высшую зону слухового восприятия, подтвердил бы наличие клеток, посылающих нервный разряд всякий раз, когда ваши органы слуха воспринимают данную песню, независимо от инструмента или особенностей исполнения. Отмечу, что такой эксперимент никогда не проводился, это было бы слишком жестоко по отношению к человеку. Но, признавая существование универсального алгоритма функционирования неокортекса, следует признать и наличие подобных клеток. В слуховом восприятии существует точно такая же обратная связь – прогнозирование, построенное на инвариантных представлениях, – как и в зрительном. Осязание работает по тому же принципу. На людях (по вполне понятным причинам) соответствующие эксперименты никогда не проводились. В настоящее время ученые занимаются подготовкой к проведению таких опытов с использованием исследовательских инструментов, сканирующих мозг обезьяны. Сейчас, сидя за столом, я держу в руке ручку. Я могу дотронуться до ее колпачка или провести пальцами вдоль металлической скрепки, с помощью которой ручку можно закрепить в нагрудном кармане. По мере того как я проделываю эти движения разными пальцами или даже губами, последовательности сигналов, поступающие в соматосенсорную зону коры головного мозга от рецепторов осязания, постоянно меняются. Будучи столь непохожими, они попадают в разные части первичной соматосенсорной зоны. Тем не менее в зонах, высших по иерархии, наш воображаемый датчик обнаружил бы клетки, которые реагируют на ручку инвариантно. Эти клетки пребывают в активном состоянии до тех пор, пока я прикасаюсь к ручке, независимо от того, какими пальцами (или даже частями тела) я это делаю. Вдумайтесь: ваш мозг не способен распознать объект на основании единственного мгновенного сигнала. Паттерн, поступающий от ваших органов чувств в какой-то один момент времени, содержит лишь крошечную толику информации, которой недостаточно для того, чтобы определить, что же вы услышали или к чему прикоснулись. Чтобы воспринять серию слуховых паттернов (например, мелодию, слово, хлопок закрывающейся двери) или понять, что за объект у вас в руках – та же ручка, – вам необходимо время. Вы не можете узнать мелодию по одной ноте и распознать предъявленный вам объект с первого прикосновения. Следовательно, нейронная деятельность, обеспечивающая сознательное восприятие объектов окружающей действительности, должна быть более продолжительной по времени, чем каждая отдельно взятая последовательность сигналов. Таким образом, мы опять пришли к выводу: чем выше расположена зона коры головного мозга, тем меньше в ней наблюдается временных изменений. Зрительное восприятие – зависимый от фактора времени информационный поток, действующий по тому же принципу, что и звуковое восприятие или осязание. Но у зрения есть особенность, которая вносит некоторую путаницу: мы все-таки способны распознавать отдельные объекты с одной фиксации. Это умение распознавать пространственные паттерны «с первого взгляда» долгие годы вводило в заблуждение ученых, занимающихся исследованиями зрительного восприятия животных. Они, как правило, не принимали в расчет фактор времени. Действительно, в лабораторных условиях как люди, так и животные способны распознавать объекты таким образом, однако это не является нормой. Нормальное зрительное восприятие, как, например, чтение этой книги, предполагает постоянные движения глазных яблок. Интеграция разных видов сенсорного восприятияА как же ассоциативные зоны? Мы уже подробно рассмотрели закономерности прохождения информационных потоков в пределах одной сенсорной зоны коры головного мозга. Сигналы обратной связи дополняют текущие сигналы и дают возможность прогнозировать будущие события. Подобные процессы происходят перекрестно и между разными сенсорными каналами восприятия, т. е. между зрением, слухом и осязанием. Например, на основе услышанного я составляю прогноз о том, что сейчас увижу или почувствую. Сейчас, печатая эти строки, я нахожусь в своей спальне. На ошейнике моей кошки Кео есть бубенчик, который позванивает, когда она ходит. И вот я слышу приближающийся звук бубенчика. На основании этого звукового сигнала я догадываюсь, что сейчас увижу свою кошку. И действительно, повернув голову в направлении двери, я вижу Кео. Я ожидал ее увидеть на основе услышанных звуков. Если бы Кео не вошла или вошло бы какое-то другое животное, я бы очень удивился. В приведенном примере звуковой сигнал вызвал звуковое распознавание Кео. Информационный поток прошел все уровни иерархии слуховых зон неокортекса и поступил в ассоциативную зону, связывающую слуховое и зрительное восприятие. На основании инвариантного представления сформировался прогноз, который по каналу обратной связи поступил назад – в зрительную и слуховую зоны. Этот процесс схематически изображен на рис. 6.4. Мозг постоянно продуцирует полисенсорные прогнозы такого типа. Я отгибаю скрепку на ручке и ожидаю, что, стоит мне отпустить пальцы, прозвучит характерный щелчок, вызванный ударом скрепки по корпусу ручки. Не услышав этого щелчка, я бы очень удивился. Мой мозг точно прогнозирует, когда я услышу звук и каким именно он будет. Для того чтобы такой прогноз осуществился, информация пересекает по иерархии все соматосенсорные зоны коры головного мозга и по обратной связи поступает назад по иерархии и слуховой зон. В результате описанного процесса возникает ожидание услышать и почувствовать щелчок. Приведу другой пример: несколько раз в неделю я добираюсь на работу велосипедом. В такие дни я иду утром в гараж, беру свой велосипед, разворачиваю его и вывожу через калитку. Пока я проделываю все указанные манипуляции, в мой мозг поступает множество зрительных и слуховых сигналов. Ежесекундно формируются полисенсорные прогнозы – предположения неокортекса относительно того, что я услышу и почувствую в следующее мгновение. Я вижу, как велосипед переезжает порог, и это вызывает у меня ожидание характерного звука. Задев ногой педаль, я наклоняю голову и ожидаю увидеть ее именно около моей ноги. Прогнозы настолько точны, что в случае возникновения малейших отклонений от ожидаемого сценария развития событий я бы тотчас заметил эти отклонения. Информационный поток движется в сенсорной иерархии одновременно в двух направлениях, таким образом формируется унифицированное восприятие, неотъемлемой частью которого является прогнозирование. 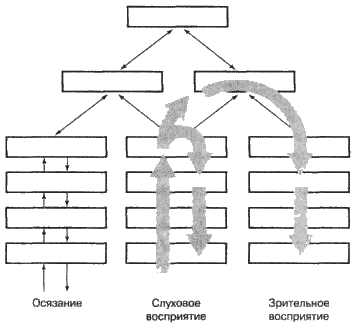 Рис. 6.4. Потоки информационных сигналов передвигаются вверх по иерархически организованным сенсорным зонам, а затем возвращаются к низшим зонам. Таким образом формируется объединенный сенсорный опыт и осуществляется прогностическая функция Предлагаю вам провести следующий опыт: оторвитесь от чтения и что-нибудь сделайте. Что угодно, лишь бы вы двигались и манипулировали каким-то предметом. Можете, например, подойти к раковине и повернуть ручку крана. Делая это, попробуйте обратить внимание на каждый звук, каждое осязательное ощущение, каждое изменение визуальной «картинки». Будьте внимательны, помните, что каждое движение тесно связано с созерцанием, слухом и осязательными ощущениями. Покрутите ручку крана вперед-назад. Ваш мозг ожидает почувствовать давление на кожу и сопротивление ваших мускулов. Вы ожидаете увидеть и почувствовать прокручивание крана, вы ожидаете увидеть и почувствовать воду. Когда вода долетит до раковины, вы ожидаете услышать другой звук – звук падающей воды, увидеть и почувствовать на своей коже обжигающе горячие (или ледяные – в зависимости от того, какой кран включаете) брызги. При ходьбе вы всегда – осознанно или нет – ожидаете услышать звук каждого шага. Даже то, что вы просто держите эту книгу в руках, тоже вызывает множество сенсорных прогнозов. Представьте себе, что вы почувствовали, будто книга закрывается, хотя зрение подсказывает вам совершенно другое: книга остается раскрытой. Наверняка вы испытали бы полное замешательство, вплоть до шока. Помните наш воображаемый эксперимент с изменением двери, описанный в главе 5? Тогда мы убедились, что мозг постоянно занят прогнозированием, причем это касается абсолютно всех сенсорных модальностей. Каждый раз, сосредоточиваясь на всех своих текущих ощущениях, вплоть до самых незначительных, я просто поражаюсь, насколько интегрированными являются все наши перцепционные прогнозы. Хотя они могут казаться простыми, даже тривиальными, надо помнить, насколько они всеобъемлющи. Они могут формироваться только на основе мощных скоординированных потоков информации, непрерывно циркулирующих в двух противоположных направлениях по иерархии зон коры головного мозга. Осознав столь тесную взаимосвязь чувств, мы делаем вывод, что вся кора головного мозга, все ее сенсорные и ассоциативные зоны действуют как единое целое. Да, у нас есть зрительная область коры, но она – лишь часть единой всеобъемлющей сенсорной системы, объединяющей восприятие изображения, звуков, прикосновений и многое другое. И в этой системе информационные потоки циркулируют вверх и вниз по сложной разветвленной иерархии. Следует также отметить, что в основе всех наших предположений лежит непосредственный опыт. Я ожидаю, что отведенная скрепка ручки при отпускании ударится о корпус ручки и издаст характерный звук только потому, что так случалось в прошлом. Звук велосипеда предсказуем по той причине, что я вывожу его из гаража далеко не в первый раз. Ни один бит подобной информации не дается нам от рождения. Все эти сведения сохраняются в мозге только благодаря его невероятно огромной способности запоминать входящую информацию. Если среди информации, поступающей в мозг, встречаются постоянные последовательности сигналов, неокортекс сохраняет их, а в будущем использует для прогнозирования событий. Хотя на рис. 6.3 и 6.4 не изображена моторная зона коры головного мозга, у нее такая же иерархическая структура, как и у описанных выше сенсорных зон. Ее можно представить как подобную сенсорную систему, но соединенную с органами чувств посредством ассоциативных зон (только с более тонкими связями с соматосенсорной зоной, что обеспечивает способность двигаться). Таким образом, моторная зона коры головного мозга функционирует подобно сенсорным зонам; кроме того, они активно взаимодействуют. Сигнал, поступающий от любого сенсорного анализатора, перемещается вверх к ассоциативной зоне, что вызывает перемещение другого импульса вниз, к моторной зоне коры головного мозга, в результате чего возникает определенное поведение. Подобно тому, как зрительный сигнал может вызвать поступление импульса в осязательную и слуховую зоны, он также может активизировать моторную зону коры головного мозга. В первом случае мы интерпретируем обратные сигналы как прогнозы, во втором – как двигательные команды. Как заметил Маунткастл, моторные зоны коры устроены так же, как и сенсорные. Кора генерирует сенсорные прогнозы так же, как моторные команды. Вскоре мы убедимся, что в неокортексе нет «чисто сенсорных» и «чисто моторных» зон. Сенсорные потоки одновременно поступают отовсюду и, возвращаясь потом по иерархии зон, формируют прогнозы или генерируют моторные команды. Хотя у моторной зоны коры головного мозга есть свои особенности, вполне корректно считать ее частью общей иерархической запоминающе-прогностической системы. Ее можно считать еще одним органом чувств. Зрение, осязание, слух и действие тесно переплетены друг с другом. Новый взгляд на зону V1Продолжая разбираться в запутанной архитектуре коры головного мозга, попробуем по-новому взглянуть на зоны неокортекса. Нам уже известно, что высшие зоны коры головного мозга формируют инвариантные представления, но почему столь важная функция должна осуществляться только в высших зонах? Не забывая о понятии симметрии, сформулированном Маунткастлом, я занялся изучением различных способов связи между зонами коры головного мозга. На рис. 6.1 показаны четыре зоны зрительного восприятия: V1, V2, V4 и IT. Как правило, каждая из них считается отдельной и непрерывной: все клетки зоны V1, обрабатывая каждая свою область зрительного поля, делают одно и то же, все клетки зоны V2 выполняют одинаковые задачи, все клетки из V4 одинаково специализированы. Как же выглядит распознавание лица с точки зрения традиционного подхода? Когда изображение лица фиксируется клетками, образующими зону V1, вначале создается грубый «эскиз» лица, состоящий из линий и других простейших элементов. Затем «набросок» поступает в зону V2, где происходит более сложный анализ черт лица. Модифицированная информация передается в зону V4 и так далее. Распознавание объекта осуществляется только в зоне IT, которой свойственна инвариантность представлений. К сожалению, при таком подходе возникает масса вопросов. Почему инвариантные репрезентации действуют только в зоне IT? Если все зоны коры головного мозга выполняют сходные функции, чем именно примечательна эта? И еще: лицо может быть воспринято правой или левой частью зоны V1, и в обоих случаях мозг его распознает. В то же время экспериментальным путем было установлено, что между этими частями зоны V1 нет непосредственного взаимодействия. Левая часть зоны V1 не знает, что видит правая. Давайте подумаем, какой вывод можно сделать. Судя по всему, разные части зоны V1 выполняют сходные функции, поскольку участвуют в распознавании лица. И в то же время физически они независимы друг от друга. Подзоны, или кластеры зоны V1, физически не связаны, однако выполняют одну и ту же функцию. И наконец, результаты исследований показывают, что все высшие зоны коры головного мозга получают объединенные сигналы от двух и более сенсорных зон, расположенных ниже в иерархии (рис. 6.3). Данный рисунок является схематическим, в действительности каждая ассоциативная зона может получать сигналы из десятка и более низших зон. Согласно традиционным представлениям, взаимосвязь между низшими зонами (например, V1, V2, V4) иная. Каждая из них якобы имеет единственный источник входящей информации, единственный поток, поступающий из низшей зоны, который не имеет отношения к потокам информации, поступающим из других зон. (Например, принято считать, что зона V2 получает информационный поток от V1, и ее работа состоит только в этом.) Но почему тогда одни зоны коры головного мозга получают объединенную информацию, а другие – нет? Это еще одно несоответствие с гипотезой универсального алгоритма неокортекса, сформулированной Маунткастлом. По этой и другим причинам я пришел к выводу, что не следует рассматривать области V1, V2, V4 как цельные зоны. Каждая из них образована более мелкими подзонами. Давайте вернемся к воображаемой обеденной салфетке (аналогии с распластанной корой головного мозга). Допустим, мы решили отметить ручкой различные функциональные зоны. Самой большой будет зона V1 – первичная зрительная зона. Следующей по размеру – зона V2. По сравнению со всеми остальными зонами эти две просто огромны. Я веду к тому, что V1 нужно рассматривать как эквивалент многих малых зон. Вместо того чтобы обвести большую зону на нашей салфетке, мы нарисуем множество маленьких зон, которые и займут территорию, принадлежащую V1. Другими словами, V1 состоит из многих малых зон коры головного мозга, которые связаны друг с другом лишь опосредованно, через более высокие зоны иерархии. В V1 входит наибольшее количество подзон по сравнению с другими зонами зрительного восприятия. V2 состоит из меньшего количества зон, но они имеют большие размеры, чем подзоны V1. То же самое утверждение действительно и для V4. Вот когда вы доберетесь до IT, она действительно окажется однородной и унифицированной. Этим объясняется тот факт, что именно клетки зоны IT формируют общую картину всего визуального восприятия окружающего мира. 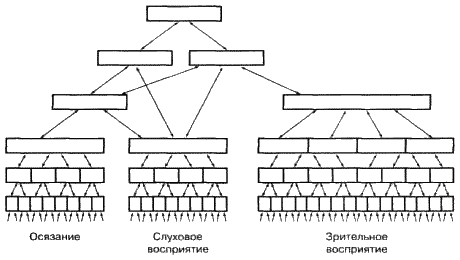 Рис. 6.5. Альтернативный подход к рассмотрению иерархии коры головного мозга Взгляните на рис. 6.5. Не правда ли, занятная симметрия? Здесь изображена та же иерархия, что и на рис. 6.3, но с учетом только что приведенной информации. Обратите внимание, что кора головного мозга везде устроена одинаково. Любая из зон получает информацию от многих других зон низшего уровня. Эта же получающая зона направляет вниз по иерархии сигналы о предполагаемых событиях. Высшие ассоциативные центры интегрируют информацию, полученную от нескольких органов чувств, например, зрительную и осязательную. Зоны низшего уровня, например, подзоны, входящие в зону V2, интегрируют информацию, воспринимаемую несколькими отдельными подзонами, входящими в V1. Зона в целом не интерпретирует – она и не может этого делать – каждый из отдельно взятых сигналов. Подзоне, входящей в V2, совершенно необязательно знать, что она получает входные сигналы из нескольких частей зоны V1. Ассоциативной зоне совершенно необязательно различать входящие в нее сигналы от зрительного и слухового анализаторов. Задача каждой зоны коры головного мозга состоит, скорее, в том, чтобы выяснить, как связаны между собой входящие сигналы, запомнить последовательность взаимосвязей между ними и в будущем использовать эту информацию для прогнозирования. Кора головного мозга всегда остается корой головного мозга. Это универсальный алгоритм функционирования неокортекса: во всех зонах происходят одни и те же процессы. Новая иерархическая схема поможет нам понять, как формируются инвариантные представления. На низшем уровне обработки визуальных сигналов левая часть зрительного поля отличается от правой части зрительного пространства так же сильно, как зрительное восприятие отличается от слухового. Левая и правая части зоны V1 формируют похожие репрезентации только потому, что получили сходный предшествующий «жизненный опыт». Их можно рассматривать как два независимых сенсорных потока, такие же, как слуховое и зрительное восприятие, которые объединяются в зонах более высокого порядка. Подобно этому, более мелкие зоны в составе V2 и V4 являются ассоциативными зонами зрительного восприятия. (Подзоны могут пересекаться, но это не меняет функционирование зон радикально.) Рассмотрение коры головного мозга под таким углом никоим образом не противоречит нашим знаниям о ее анатомическом строении. Информационные потоки передвигаются по всем восходящим и нисходящим ветвям иерархического древа системы памяти. Сигнал, поступивший в левую часть зрительного поля, может привести к формированию прогнозов для правой части зрительного поля, как звук колокольчика на шее кошки стал причиной визуального прогноза о том, что она войдет в спальню. Главная польза нашего нового схематического изображения заключается в том, что теперь мы можем быть уверены: в формировании инвариантных репрезентаций участвует каждая зона коры головного мозга. Согласно традиционным взглядам, о полных инвариантных представлениях речь не шла до тех пор, пока сигнал не попадал в высшие зоны неокортекса (такую, например, как зрительная зона IT, в которой формируется представление об общей наблюдаемой визуальной картине). Теперь мы установили, что на самом деле инвариантные репрезентации вездесущи – они формируются каждой зоной коры головного мозга. Инвариантность – не чудо, возникающее в высших зонах. Каждая зона формирует инвариантные репрезентации на основе данных, получаемых из зон, расположенных ниже по иерархической лестнице (каждая из которых «видит» лишь небольшую часть окружающего мира). Можно сказать, что уровень восприятия в низших зонах значительно беднее такового в высшей зоне IT, однако все они выполняют сходные функции: V4, V2, V1 выполняют ту же работу, что и IT. Ассоциативные зоны, расположенные выше IT, формируют инвариантные представления, основанные на информации, полученной от нескольких сенсорных анализаторов. Следовательно, каждая из зон коры головного мозга создает инвариантные представления исходя из того, что находится ниже в иерархии. Это красиво. Наша головоломка изменилась. Нам больше не нужно выяснять, каким образом за четыре шага формируются инвариантные представления. Но теперь мы должны разобраться, как формируются инвариантные репрезентации во всех зонах коры головного мозга. Если мы признаем существование универсального алгоритма коры головного мозга, то такая постановка вопроса обоснованна. Если в одной зоне хранятся последовательности сигналов, то и в остальных зонах тоже. Если одна зона создает инвариантные репрезентации, то и остальные зоны тоже. Новая схема иерархии коры головного мозга, представленная на рис. 6.5, поможет нам глубже понять суть данного подхода. Модель окружающего мираПочему кора головного мозга устроена иерархически? Вы обладаете способностью размышлять о мире, передвигаться в окружающем пространстве и прогнозировать будущее, потому что в вашем неокортексе сформирована модель внешнего мира. Одна из наиболее важных концепций данной книги состоит в том, что иерархическая структура коры головного мозга хранит модель иерархического строения внешнего мира. Вложенная структура реального мира отображается вложенной структурой коры вашего головного мозга. Что я подразумеваю под вложенной, или иерархической, структурой? Обратимся к музыке. В ней ноты объединяются в группы, которые, в свою очередь, образуют мелодические фразы. Так рождается мелодия или песня. Из песен состоят альбомы. В качестве второго, не менее удачного, примера можно привести письменность. Буквы складываются в слоги, которые сочетаются в слова. Из слов состоят предложения и фразы. Хотите еще один пример? Вспомните, что у вас расположено по соседству – дороги, школы, дома? В каждом доме есть комнаты. В каждой комнате есть стены, потолок, пол, дверь, одно или больше окон. Каждая из этих составляющих, в свою очередь, состоит из более мелких частей. Окно состоит из стекла, рамы, ручки, щеколды. Щеколда состоит из еще более мелких частей, таких как винты. Осмотритесь вокруг. Сигналы от вашей сетчатки, поступающие в первичную зону зрительного восприятия, сочетаются в линейные сегменты. Линейные сегменты объединяются в более сложные формы. Эти формы объединяются в еще более сложные образы, такие, например, как нос. Носы вкупе с глазами и ртами головной мозг объединяет в человеческие лица. Из лица, дополненного остальными частями тела, у вас складывается образ человека, сидящего напротив. Все объекты окружающего мира состоят из подобъектов, составляющих одно целое. Эти составляющие отдельных образов и являются их определениями. Называя что-либо определенным словом, мы подразумеваем, что существует набор характеристик, связанных с этим определением. Лицо называется лицом, потому что на нем всегда присутствуют глаза, рот, нос. Глаз является глазом, потому что в нем всегда присутствуют зрачок, радужная оболочка, ресницы, веки и так далее. То же самое можно сказать и о стульях, машинах, деревьях, парках, странах. Песня называется песней благодаря определенной последовательности звуков. Весь мир, окружающий нас, – это своего рода песня. Каждый объект в нем состоит из множества более мелких, а большинство малых объектов являются частью больших. Именно это я и подразумеваю под вложенной структурой, характерной практически для всех объектов окружающей среды. Аналогичным образом, информация об объектах и способы их представления сохраняются в иерархической структуре коры головного мозга. Информация о том, что такое здание, хранится не в единственной зоне неокортекса, а в иерархии зон. Соответственно, эта информация отражает иерархическую структуру понятия «здание». Фундаментальные взаимосвязи сохраняются в верхней части иерархии зон, а более специфичные – ближе к «подножию» иерархической лестницы. Иерархические связи внешнего мира подтверждаются в ходе процесса обучения. Никому из нас не даются от природы знание языка, образов окружающего мира, музыки. Но в кору нашего головного мозга встроен разумный самообучающийся алгоритм, который естественным образом выявляет иерархические структуры и запоминает их. Если никакой структуры нет, мы испытываем смятение и считаем сложившуюся ситуацию хаотичной. В каждый отдельно взятый момент времени вы можете воспринимать только определенную часть внешнего мира. Вы можете находиться только в одной комнате вашего дома и смотреть только в одном направлении. Благодаря иерархическому строению коры головного мозга вы в состоянии осознавать, что находитесь дома, в своей гостиной, и что вы смотрите в окно, даже если в данный момент ваш взгляд остановился на щеколде. Высшие зоны коры головного мозга поддерживают инвариантную репрезентацию вашего дома, более низкие – репрезентацию комнаты, а первичные – анализируют непосредственные визуальные символы, в данном случае – происходящее за окном. Точно так же иерархическое строение неокортекса позволяет вам осознавать, что вы прослушиваете отдельную песню из определенного музыкального альбома, несмотря на то что в каждый отдельно взятый момент вы слышите только одну ноту, которая сама по себе ничего для вас не значит. Благодаря иерархическому строению неокортекса вы понимаете, что беседуете со своим лучшим другом, хотя в данный конкретный момент вы смотрите только на его ладонь. Высшие зоны коры головного мозга выполняют стратегическую функцию – следят за ходом развития событий в крупных масштабах, в то время как более низкие зоны сосредоточены на восприятии скоротечных аспектов каждой конкретной ситуации. Поскольку в некий отдельно взятый момент времени вы можете видеть, слышать и ощущать только очень небольшую часть внешнего мира, информация поступает в мозг в виде последовательностей сигналов. Кора головного мозга особенно чувствительна к тем последовательностям, которые постоянно повторяются, и стремится их запомнить. В некоторых случаях, таких как, например, звучание мелодии, последовательность сигналов, поступающая в ваш головной мозг, определенным образом организована. Надеюсь, что концепция последовательностей сигналов понятна любому из вас. Но я собираюсь использовать термин последовательность в более широком смысле, приближающемся к математическому термину ряд. Последовательность – это ряд сигналов, которые следуют друг за другом, но не всегда в строгом порядке. Для большей наглядности предлагаю вам рассмотреть примеры. Когда я смотрю на ваше лицо, последовательность входных сигналов, поступающих в кору головного мозга, не остается неизменной, она определяется саккадами моих глаз. Один раз мой взгляд движется по пути «глаз-глаз-нос-рот», а через мгновение порядок изменяется: «рот-глаз-нос-глаз». Части лица – это последовательности. Они статистически связаны и встречаются близко во времени, хотя их порядок может меняться. Если вы воспринимаете «лицо», фиксируя взгляд на «носу», наиболее вероятным следующим паттерном будет «глаз» или «рот», а не «ручка» или «машина». В каждую область коры поступает поток таких паттернов. Если зона коры может предсказать, какой из паттернов последовательности будет следующим, она формирует постоянное представление этой последовательности – запоминает ее. Запоминание последовательностей это самый главный компонент формирования инвариантных репрезентаций объектов реального мира. Эти объекты могут быть конкретными, как ящерица, лицо или дверь, или абстрактными, как слово или теория. Мозг рассматривает абстрактные и конкретные объекты одинаково. И те, и другие – всего лишь предсказуемые последовательности паттернов, которые время от времени повторяются. Именно благодаря повторяемости зона коры головного мозга определяет, что она имеет дело с объектом реального мира. Предсказуемость – это самое сердце реальности. Зона коры может обнаружить, что во время физических движений (таких как саккады глаз или движения пальцев) паттерны гарантированно и предсказуемо сменяют друг друга, или же она может их предсказывать по мере того, как они разворачиваются во времени (как, например, звуки, из которых состоит слово или мелодия). В этом случае мозг решает, что здесь есть причинная связь. Вероятность того, что многочисленные паттерны будут неоднократно встречаться вместе чисто случайно, ничтожно мала. Предсказуемая последовательность паттернов должна быть частью реального объекта, который существует на самом деле. Поэтому возможность надежно предсказывать – «железный» способ узнать, что какие-то события в мире на самом деле связаны. Нос, глаза, уши и рот есть у любого лица. Если мозг воспринимает глаз, после саккады – еще один, после следующей саккады – рот, значит, он точно имеет дело с лицом. Если бы зоны коры могли говорить, мы бы услышали что-то вроде: «Я воспринимаю много разных паттернов. Иногда я не могу предсказать, какой из них будет следующим. Но между ними точно есть связь. Я всегда наблюдаю их вместе и уверенно могу перескакивать от одного к другому. Поэтому я дала этим событиям общее имя и употребляю его всякий раз, когда мне попадается любое из них. Именно это групповое имя, а не разрозненные паттерны, я передаю высшим зонам коры». Следовательно, можно сказать, что мозг хранит последовательности последовательностей. Каждая зона коры выделяет последовательности, дает им то, что я называю «именами», и передает эти имена зонам, которые расположены выше в иерархии. Последовательности последовательностейМы видим все меньше и меньше временных изменений информации по мере того, как она поднимается по иерархии зон коры головного мозга. В первичных визуальных зонах вроде V1 совокупность активных клеток быстро изменяется по мере того, как несколько раз в секунду на сетчатку поступают новые паттерны. А вот в визуальной зоне IT возбужденные клетки более стабильны. Что же происходит? Каждая область коры владеет определенным репертуаром последовательностей, подобным репертуару певца. Последовательности-песни могут быть о чем угодно: как шумит прибой, как выглядит лицо вашей матери, как пройти от вашего дома к магазину на углу, как произносится слово «попкорн», как тасовать колоду карт. Мы даем имена песням, и каждая зона коры тоже дает имена всем последовательностям, которые ей известны. Это «имя» – группа клеток, чья коллективная активность обозначает объекты последовательности (пока не думайте о том, как выбирают клетки для того, чтобы обозначать последовательность, – с этим мы разберемся чуть позже). Означенные клетки будут активны, пока длится последовательность, и именно это «имя» передается вверх по иерархии. До пор пока входные сигналы являются частью прогнозируемой последовательности, зона коры головного мозга посылает к высшим зонам одно и то же «имя». Зона, принимающая входные сигналы, словно отправляет наверх сообщение: «Вот имя последовательности, которую я вижу, слышу или осязаю. Отдельные ноты, контуры или качество поверхности не имеют значения. Я обязательно вам сообщу, если произойдет что-то новое или непредвиденное». Так, например, высшая зона зрительной иерархии сообщает ассоциативной зоне: «Я вижу лицо. Каждая саккада дает фиксацию на различных частях лица. Я вижу различные части лица в определенной последовательности, но это точно одно и то же лицо. Я вам сообщу, если встретится что-то новое». Таким образом, предсказуемой последовательности событий соответствует «имя» – постоянный паттерн импульсов от возбужденных клеток. Продвигаясь вверх по иерархической пирамиде, мы будем видеть это снова и снова. Одна зона может заниматься распознаванием звуков, из которых состоят фонемы: она посылает сигнал, представляющий фонемы, – в более высокую зону иерархии. Следующая зона распознает последовательность фонем и формирует из них слова. Следующая, более высокая, зона распознает слова и формирует из них фразы и так далее. Не забывайте, что в более низких зонах «последовательность» может быть относительно простой, например, кромка чего-либо, передвигающаяся в пространстве. Давая имена прогнозируемой последовательности сигналов в каждой зоне иерархии, мы получаем все более и более высокую стабильность наверху иерархии. Так создаются инвариантные репрезентации. Обратный процесс происходит, когда сигнал перемещается по обратной связи вниз по иерархии: устойчивые репрезентации «разворачиваются» в последовательности сигналов. Предположим, вы решили вспомнить содержание Геттисбергской речи Линкольна, которую учили в седьмом классе. В одной из верхних зон коры вашего головного мозга хранится модель, которая является репрезентацией известной речи Линкольна. Сначала модель разветвляется на последовательности фраз. Зоной ниже каждая фраза разворачивается в последовательности слов. На этом этапе сигнал расщепляется и движется в зрительную и моторную зоны коры головного мозга. Спускаясь по моторному пути, каждое слово расщепляется на усвоенную последовательность фонем. В конце концов, на самом низком уровне, каждая фонема расщепляется на последовательность артикуляционных команд, обеспечивающих произнесение звуков. Чем ниже по иерархии мы спускаемся, тем быстрее меняются сигналы. Одна цельная репрезентация, хранящаяся в высшей зоне иерархии, преобразуется в продолжительную и сложную последовательность звуков речи. По мере того как информационный поток спускается вниз по иерархии, инвариантность приобретает особое значение. Если вы хотите напечатать Геттисбергскую речь, не произнося ее вслух, то начнете с той же самой репрезентации из высшей зоны. В следующей зоне сигнал разделится на фразы, потом – на слова. До этого момента никаких различий не наблюдается. Но далее моторная зона коры головного мозга избирает иной путь. Слова расщепляются на буквы, а затем формируются мышечные команды, поступающие к вашим пальцам с тем, чтобы вы начали печатать. Итак, усвоенная вами речь Линкольна представлена в головном мозге в виде инвариантной репрезентации. Нет никакой разницы в том, будете ли вы ее произносить, печатать на компьютере или записывать от руки. Обратите внимание: вам не нужно запоминать речь дважды (раз для письменного воспроизведения и второй – для устного произношения). Единожды усвоенная информация о речи Линкольна может привести к различным поведенческим проявлениям. В любой зоне коры головного мозга инвариантный сигнал может разветвляться и передвигаться вниз по иерархии различными путями. Как еще одно проявление эффективной работы головного мозга, репрезентации простых объектов из нижней части иерархии корковых зон могут использоваться повторно для формирования разных последовательностей сигналов в высших зонах. Например, нам не нужно запоминать один набор слов и фонем для воспроизведения вслух Геттисбергского обращения и совершенно другой набор – для речи Мартина Лютера Кинга «У меня есть мечта», хотя эти два выступления имеют некоторые общие слова. Иерархия вложенных последовательностей обеспечивает повторное использование объектов более низкого уровня (например, слов, фонем, букв). Это исключительно эффективный способ хранения информации о внешнем мире и его структуре, и он очень сильно отличается от принципов работы компьютера. В сенсорных областях происходит такое же разворачивание последовательностей, как и в моторных зонах. Этот процесс позволяет вам ощущать и воспринимать все многообразие объектов окружающей среды. Когда вы подходите к холодильнику за порцией мороженого, зрительная зона коры головного мозга активизируется на многих уровнях. На высшем уровне вы постоянно «видите» холодильник. На более низких уровнях зрительное ожидание подразделяется на серию более специфических, локализированных на отдельных объектах зрительных сигналов. Зрительное восприятие холодильника складывается из фиксаций взгляда на ручке дверцы, самой дверце, прикрепленных магнитах, детских рисунках и так далее. За несколько миллисекунд, которые проходят в период саккады от одного свойства холодильника к другому, прогнозы результатов каждой саккады устремляются вниз по зрительной иерархии. До тех пор пока прогнозы от саккады до саккады оправдываются, высшие зоны зрительного восприятия твердо уверены в том, что вы действительно видите перед собой свой холодильник. Обратите внимание, что в отличие от строгого порядка слов в Геттисбергском обращении, та последовательность паттернов, с которой вы имеете дело в случае с холодильником, может оказаться изменчивой. Поток входных сигналов и прогнозы о них зависят от ваших действий. Значит, в случаях, подобных этому, разветвляющиеся сигналы не являются стабильной последовательностью, но по сути своей процесс все равно остается одним и тем же: стабильные, мало меняющиеся репрезентации высшего уровня, которые разветвляются в быстро меняющиеся паттерны низших корковых зон. Тот способ, которым мы запоминаем последовательности и представляем их под каким-либо именем во время передвижения информационных потоков вниз и вверх по иерархии коры головного мозга, чем-то напоминает иерархию армейских команд. Генерал отдает приказ: «На зиму перебросьте военные части во Флориду». Простая команда высшего уровня по мере ее передвижения вниз по армейской иерархии расщепляется на все более детализированные последовательности команд. Подчиненные генерала знают, что команда требует определенной последовательности шагов, таких как приготовления к передислокации, транспортировка во Флориду, подготовка к обустройству на новом месте. Каждый из этих шагов разбивается, в свою очередь, на еще более специфические задания, которые адресуются подчиненным. На нижней ступени иерархии будут стоять тысячи рядовых солдат, предпринимающих десятки тысяч физических шагов для того, чтобы военные части были переведены во Флориду. Что касается обратной связи, то на каждом уровне формируются отчеты о проделанной работе. В результате генерал получает доклад: «Передислокация во Флориду осуществлена успешно», но при этом не вникает во все подробности инициированного им процесса. У данного правила есть исключение. Если возникает проблема, которая не может быть разрешена подчиненными на соответствующем низком уровне, запрос передается к более высоким уровням иерархии до тех пор, пока решение не будет найдено. Но офицер, который знает, как решить возникшую проблему, не рассматривает ее как исключение. То, что является неожиданной проблемой для его подчиненных, для него самого является следующим по списку заданием – он лишь отдает подчиненным новые команды. Кора головного мозга функционирует по аналогичному принципу. Когда происходит неожиданное событие (поступают непрогнозированные сигналы), информация передается в более высокие по иерархии зоны коры головного мозга до тех пор, пока возникшая трудность будет разрешена. Если более низкие зоны не в состоянии спрогнозировать поступающие сигналы, они классифицируют их как ошибку и передают в высшие «командные центры». Процедура повторяется до тех пор, пока следующая зона не сможет распознать поступающий сигнал. Каждая зона коры головного мозга устроена так, что старается сохранять и активизировать последовательности сигналов. Однако такое представление о мозге является слишком упрощенным. Давайте немного усложним нашу модель. Сигналы поступают в зоны коры головного мозга по тысячам или даже миллионам аксонов. Эти аксоны идут от самых разных областей и по ним передаются самые разные последовательности сигналов. Потенциальное количество сигналов на одной тысяче аксонов превышает количество молекул во Вселенной. Каждая зона коры головного мозга на протяжении всего времени своего существования воспринимает лишь небольшую часть этого невероятного количества сигналов. Возникает вопрос: из чего состоят последовательности сигналов, сохраняющиеся в различных зонах неокортекса? Я предполагаю, что сначала зона классифицирует входные сигналы на основе ограниченного числа возможных вариантов, а потом формирует последовательности. Представьте себе, что вы – зона коры головного мозга. Ваша задача состоит в том, чтобы рассортировать цветные листы бумаги. У вас есть десять ведер, каждое из которых помечено определенным цветом. Одно ведро предназначено для листов зеленого цвета, другое – для красных, следующее – для желтых и так далее. Каждый из предлагаемых вам листов немного отличается по цвету. Поскольку количество существующих в мире цветов не ограничено, то не найдется даже двух листов бумаги одинакового цвета. Иногда очень легко определить, в какое из ведерок нужно отправить лист, а порой наоборот – очень трудно. Лист, окрашенный в промежуточный между красным и оранжевым цвет, можно бросить и в то, и в другое ведро, но вам нужно выбрать только одно, даже если вы сделаете это наугад. Задача данного упражнения состоит в том, чтобы показать, как мозг должен классифицировать сигналы. Зоны коры головного мозга именно этим и занимаются, с той лишь разницей, что у них не имеется контейнеров, в которые они бы помещали сигналы. А сейчас вам выпадает шанс найти последовательность. Вы замечаете, что листы очень часто следуют в таком порядке: красный, красный, пурпурный, оранжевый, зеленый. Вы присваиваете этой последовательности имя «ккпоз». Обратите внимание, что распознавание какой бы то ни было последовательности было бы невозможным, если бы вы сначала не рассортировали все листы по десяти ведрам. Не рассортировав листы по десяти из возможных категорий, вы бы не смогли определить, что две последовательности одинаковы. Сейчас вы собираетесь проанализировать все входные сигналы – цветные листы бумаги, соответствующие в нашей аналогии сигналам, получаемым из низших зон коры головного мозга. Вы сортируете их и пытаетесь обнаружить последовательности. Оба шага – сортировка и формирование последовательностей – являются необходимыми для создания инвариантных репрезентаций, и любая зона коры головного мозга именно этим и занимается. Процесс формирования последовательностей окупается с лихвой, когда входные сигналы неоднозначны, как, например, лист бумаги, который можно назвать и красным, и оранжевым. Допустим, вам предстоит определиться, какое ведро выбрать для этого листа, даже если вы не уверены наверняка в своем выборе. Зная наиболее вероятную последовательность для серии входных сигналов, вы сможете использовать эти знания для классификации неоднозначных входных сигналов. Если вы считаете, что попали на «ккпоз»-последовательность, поскольку вам попались два красных, зеленый и пурпурный цвета, то у вас формируется ожидание, что следующий цвет будет оранжевым. Но оказывается, что следующий лист совсем не оранжевый, он, скорее, нечто между красным и оранжевым. Вполне может быть, что он даже более близок к красному цвету, чем к оранжевому. Вы же ожидали «ккпоз»-последовательность, вы знакомы с ней, поэтому положите лист в ведро, предназначенное для оранжевых листов. В сомнительных случаях вы наверняка воспользуетесь контекстом известных последовательностей. Этот феномен постоянно подтверждается в повседневной жизни. Когда люди разговаривают, то часто «глотают» отдельные слова, понять которые вне контекста было бы невозможно. Тем не менее такая особенность устной речи не мешает собеседникам прекрасно понимать друг друга. Бывает, невозможно прочесть отдельно взятое написанное от руки слово, смысл которого проясняется при прочтении целой фразы. В подавляющем числе таких случаев вы не осознаете, что восполняете недостающую информацию, исходя из усвоенных ранее последовательностей сигналов. Вы слышите то, что ожидаете услышать, и видите то, что ожидаете увидеть, тогда, когда услышанное и увиденное вами соответствует прошлому опыту. Обратите внимание, что запоминание последовательностей не только облегчает интерпретацию неоднозначных входных сигналов, но и позволяет спрогнозировать, какой сигнал окажется следующим. Высшие зоны коры головного мозга сообщают зонам более низкого порядка, чего им следует ожидать дальше. Возвращаясь к примеру с сортировкой бумаги, вы могли бы сказать человеку, передающему вам листы: «Послушай, если ты не можешь никак решить, какого цвета лист подавать мне следующим, то, насколько я помню, это должен быть оранжевый». Распознавая последовательности сигналов, вышестоящая зона коры предсказывает, каким будет следующий сигнал, и сообщает зонам нижестоящим, чего им следует ожидать. Зона коры головного мозга не только усваивает определенные последовательности, но и модифицирует уже имеющиеся классификации. Допустим, вы начали заполнять ведра зеленого, желтого, красного, пурпурного и оранжевого цветов. Вы уже приготовились к поступлению листов в последовательности «ккпоз» (или похожей комбинации этих цветов). Но что, если ваши ожидания не оправдаются? Что, если всякий раз, когда вы столкнетесь с подобием данной последовательности, вместо пурпурного вам предоставят лист цвета индиго? Скорее всего, вы измените надпись на соответствующем ведре с «Пурпурного» на «Индиго». Так вы приспосабливаетесь к меняющимся обстоятельствам и уходите от двусмысленности. И все это благодаря поразительной гибкости неокортекса. Подобные изменения уже сложившихся классификаций происходят в зонах коры головного мозга на протяжении всей вашей жизни. Данный процесс осуществляется на основе взаимодействия последовательностей сигналов; он же положен и в основу обучения. Все зоны коры головного мозга очень пластичны, т. е. информация, сохраняющаяся в них, модифицируется под влиянием прошлого опыта и восприятия текущей ситуации. Создавая новые классификации и новые последовательности, вы запоминаете мир. И наконец, давайте обсудим, как эти классификации и прогнозы взаимодействуют с высшими зонами коры головного мозга. Здесь следует представить вам еще одну функцию коры головного мозга – трансляцию воспринятого в зону на уровень выше (в нашем случае это будет передача вверх по иерархии листа с отметкой «ккпоз»). Для следующего уровня буквы сами по себе мало что означают. Название – это просто сигнал, подлежащий последующей обработке и сопоставлению с остальными сигналами, классификации и последующему размещению в последовательности более высокого порядка. Точно так же, как и вы, эта высшая зона следит за последовательностями, которые воспринимает. В какой-то момент она может обратиться к вам: «Эй, послушайте, если вы не знаете, что мне подавать следующим, то, насколько мне известно, это должна быть последовательность „жжкзж“». Фактически, это прямое указание на то, что вы должны искать в данный момент в своем потоке входящих сигналов. Вы будете изо всех сил стараться распознать составляющие компоненты известной вам последовательности. Поскольку многие из нас сталкивались с понятием классификации сигналов, принятым в теории искусственного интеллекта и используемым при описании зрительного восприятия, давайте рассмотрим, чем отличаются эти сигналы от функционирования коры головного мозга. Пытаясь обучить компьютер распознавать объекты, исследователи, как правило, создают шаблон, например, изображение чашки или какую-то прототипную форму чашки, а потом программируют машину на поиск соответствия входящей информации шаблону. В случае значительного сходства выдается ответ, что обнаружена чашка. Отличие реального интеллекта состоит в том, что в нашем мозге нет таких шаблонов, а сигналы, поступающие в верхние зоны коры головного мозга, совсем не похожи на картинки. Вы не запоминаете моментальных изображений, воспринимаемых вашей сетчаткой, или ушами, или тактильными рецепторами, расположенными по всей поверхности вашей кожи. Иерархическое строение коры головного мозга обеспечивает распределение информации об объекте по всей иерархии – воспоминание о том или ином явлении или событии, предмете или человеке не сохраняется в каком-то одном месте. Кроме того, каждая зона из образующих иерархию формирует инвариантные запоминания, а в определенном поле неокортекса возникает последовательность инвариантных репрезентаций, которые, в свою очередь, тоже являются последовательностями инвариантных запоминаний. В головном мозге не сохраняются изображения чашки или каких-либо других объектов внешней среды. В отличие от камеры ваш мозг запоминает окружающую среду такой, какая она есть, а не такой, какой она выглядит. Когда вы думаете о мире, то вспоминаете последовательности сигналов, соответствующих свойствам объектов, их функционированию и характеристикам ощущений в определенный момент времени. Последовательности, в рамках которых вы воспринимаете объекты окружающей действительности, отражают инвариантную структуру самого мира. Порядок, в котором вы воспринимаете составляющие внешнего мира, предопределяется его структурой. Например, вы можете попасть в самолет по трапу, но не напрямую от кассы. Последовательностями, в которых вы воспринимаете мир, представлена его реалистичная структура, именно она и формируется в коре головного мозга. Не забывайте, однако, что инвариантная репрезентация в любой зоне коры головного мозга может превратиться в подробный прогноз того, какие ощущения возникнут в органах чувств, с одновременной ретрансляцией сигнала вниз по иерархии. Точно так же инвариантная репрезентация в моторной коре головного мозга может перейти в подробные ситуационно-специфические моторные команды посредством передачи сигнала вниз по иерархии. Строение зон коры головного мозгаА сейчас давайте рассмотрим отдельную зону коры головного мозга, одну из тех, что изображены блоками на рис. 6.5. 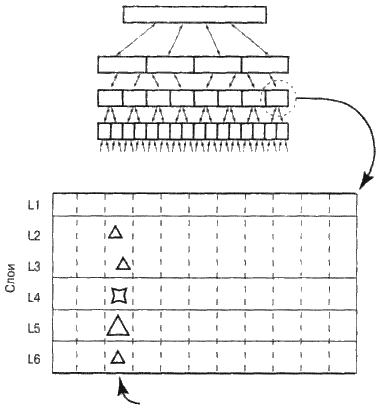 Рис. 6.6. Слои и колонки в зоне коры головного мозга На рис. 6.6 приведено схематическое строение зоны коры головного мозга. В этом разделе я покажу, как клетки зоны коры головного мозга распознают и вызывают из памяти последовательности сигналов, обеспечивая тем самым формирование инвариантных репрезентаций и составление прогнозов. Начнем с описания структуры, общей для всех зон коры головного мозга. Прежде всего, между ними имеются значительные расхождения в размерах. Наиболее обширными являются первичные сенсорные зоны. К примеру, зона V1 занимает в задней части мозга область, приблизительно соответствующую площади паспорта. Как я уже упоминал выше, она состоит из множества подзон, которые могут быть не больше букв на этой странице. Но давайте пока предположим, что типичная зона коры головного мозга размером с мелкую монету. Вспомните шесть визиток, с помощью которых в главе 3 мы представили слои ткани неокортекса. Что это за слои? Если мы рассмотрим корковую зону размером с монету под микроскопом, то увидим, что при передвижении снизу вверх плотность и форма нервных клеток различаются. Верхний слой 1 сильнее всего отличается от остальных слоев. Он состоит из густого переплетения аксонов, идущих параллельно поверхности коры, нейронов в этом слое почти нет. Строение слоев 2 и 3 сходно – они густо «населены» большим количеством пирамидных клеток. Слой 4 состоит из нервных клеток звездчатой формы. В слое 5 мы увидим как пирамидные клетки обычных размеров, так и чрезвычайно большие. Несколько типов нейронов есть и в нижнем слое 6. Визуально мы воспринимаем горизонтальные слои, но большее значение имеют колонки клеток, располагающиеся перпендикулярно. Можете считать колонки вертикальными объединениями клеток, которые работают вместе. (Термин «колонка» вызывает много дебатов в научных кругах. Нейробиологи спорят об их размере, функциях и роли в целом. Для удобства будем мыслить в рамках их архитектуры, поскольку то, что у коры столбчатая архитектура, признается всеми.) Слои в пределах каждой колонки связываются аксонами, которые проходят вверх-вниз и создают синапсы. Отмечу, что наши колонки не имеют четких очертаний – в коре нет ничего простого. То, что колонки существуют, доказывается следующими положениями. Положение 1. Вертикально расположенные клетки одной колонки активизируются под воздействием одних и тех же стимулов. Так, при подробном рассмотрении колонок зоны V1 мы обнаружим, что некоторые из них соответствуют линейным сегментам с правым уклоном (/), а другие – линейным сегментам с левым уклоном (\). Клетки из одной колонки тесно связаны, именно поэтому вся колонка реагирует на один и тот же стимул. Например, активная клетка слоя 4 активизирует расположенные над ней клетки слоев 3 и 2, что, в свою очередь, активизирует клетки, входящие в слои 5 и 6. Активность распространяется вниз и вверх в пределах колонки. Положение 2. Наличие колонок нейронов в зонах неокортекса объясняется особенностями формирования коры головного мозга в эмбриогенезе. Эмбриональные клетки-предшественники мигрируют из внутренней мозговой впадины к месту образования коры головного мозга. Каждая из указанных клеток делится, образуя приблизительно сотню нейронов, называемых микроколонками. Эти нейроны связаны друг с другом вертикально так, как я описал. Термин «колонка» часто используется достаточно свободно для описания разных явлений. Под колонкой может подразумеваться общая вертикальная связность в коре или же вполне определенная группа клеток, происходящих от одной клетки-предшественницы. В рамках последнего определения мы можем утверждать, что в коре головного мозга есть несколько сотен миллионов микроколонок. Чтобы вам легче было представить эту колончатую структуру, вообразите микроколонку в человеческий волос толщиной. Возьмите тысячи волосков и разрежьте их на небольшие сегменты, имеющие высоту строчной буквы Т. Если поставить их бок о бок очень плотно друг к другу и склеить, получится густая щетина. Теперь создайте покров из длинных и очень тонких волосков. Они будут представлять аксоны слоя 1. Приклейте их горизонтально к поверхности нашей щетины. Этот щетиноподобный коврик является упрощенной моделью зоны коры головного мозга размером с монету. Потоки информации преимущественно передвигаются преимущественно вдоль волосков: горизонтально в слое 1 и вертикально в слоях 2-6. Рассмотрим еще одну важную особенность колонок, а затем перейдем к анализу их назначения. При пристальном рассмотрении мы увидим, что не менее 90% синапсов клеток из одной колонки – это связи с клетками, расположенными вне пределов самой колонки: некоторые соединения – с соседними колонками, некоторые – вообще с другими частями мозга. На основании чего же мы говорим о важности колонок, если большинство связей распределено по всему мозгу, а не локализовано? Ответ на этот вопрос нам даст модель «Память-предсказание». В 1979 году Вернон Маунткастл не только заявил о существовании единого алгоритма коры головного мозга, но также предположил, что колонка нейронов в коре головного мозга является базовой единицей обработки информации. Правда, он не знал, какова именно функция колонки. Я считаю, что последняя является базовой единицей прогнозирования. Чтобы колонка нейронов могла прогнозировать, когда ей следует активизироваться, нервным клеткам необходимо знать, что происходит в других частях мозга. Именно в этом и заключается смысл обширных синаптических связей. Ниже мы обсудим этот интересный момент подробнее, а сейчас я хочу дать вам общее представление о назначении подобных соединений нейронов в коре головного мозга. Чтобы предсказать следующий звук мелодии, вы учитываете ее название, ту часть, которая звучит в данный момент времени, время, прошедшее с момента последнего услышанного вами звука, и то, каким он был. Большое количество синапсов, связывающих нервные клетки одной колонки с остальными частями мозга, обеспечивает контекст, в котором и осуществляется прогностическая функция коры головного мозга. Теперь обратимся к рассмотрению потоков информации, циркулирующих вверх-вниз по иерархии корковых зон. Восходящий поток сигналов проходит относительно прямой путь, который схематически отображен на рис. 6.7. Представьте, что перед нами зона коры головного мозга, состоящая из тысяч колонок. Увеличим одну из них. Входные сигналы из нижних зон попадают в слой 4, который является основным входящим слоем. По дороге они «отмечаются» в слое 6 (скоро вы поймете, почему это так важно). Клетки слоя 4 передают сигналы вверх – к клеткам слоев 2 и 3, находящимся в пределах той же колонки. Далее клетки слоев 2 и 3 передают информацию по аксонам еще выше – к входному слою следующей в иерархии зоны. Таким образом информация передвигается от зоны к зоне вверх по иерархии в пределах одной колонки нейронов. Путь нисходящего потока информации (рис. 6.8) более сложен. Клетки слоя 6 посылают информацию вниз, в слой 1 области, расположенной ниже в иерархии. Аксоны нейронов слоя 1 распространяются на очень большие расстояния. Следовательно, информация, передвигающаяся вниз, потенциально может активизировать многие колонки нейронов. Хотя «своих» клеток в слое 1 очень мало, сигналы, проходящие через этот слой, могут активизировать клетки слоев 2, 3 и 5, поскольку у последних есть дендриты в слое 1. Аксоны, выходящие из клеток слоев 2 и 3. покидая кортекс, формируют синапсы в слое 5 и, как предполагается, возбуждают клетки слоев 5 и 6. Итак, мы приходим к выводу, что информация, передвигаясь вниз по иерархии, проходит более запутанный путь, чем в случае движения вверх. Благодаря слою 1 она может распространяться в самых разных направлениях. Обратная информация генерируется в клетках слоя б высшей зоны. Затем она распространяется в слое 1 низшей зоны. Клетки слоев 2, 3 и 5 низшей зоны возбуждаются, некоторые из них, в свою очередь, возбуждают клетки слоя 6, который проецируется в слой 1 более низких в иерархии областей и так далее. (При помощи рис. 6.8 понять описанные процессы намного проще.) 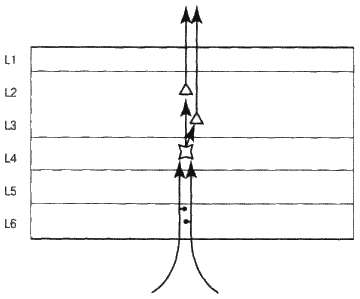 Рис. 6.7. Восходящий информационный поток Приведу предварительное объяснение того, зачем информация распределяется в слое 1. Чтобы трансформировать инвариантную репрезентацию в какой-то прогноз, ежесекундно надо решать, куда должен идти сигнал, направленный вниз по иерархии. Слой 1 обеспечивает возможность конвертирования инвариантного представления в более специфическую и конкретную репрезентацию. Например, вы можете воспроизвести Геттисбергскую речь как в устной, так и в письменной форме. Общая репрезентация трансформируется двумя разными способами: первый – для устной, а второй – для письменной формы. Горизонтальный поток активности в слое 1 обеспечивает механизм для осуществления такого процесса. Чтобы инвариантные представления высокого уровня могли транслироваться вниз и принимать форму конкретных прогнозов, мы должны располагать механизмом, позволяющим потоку сигналов разветвляться в каждом слое. На эту роль как нельзя лучше подходит слой 1. Даже не зная о его существовании, мы могли бы предвидеть, что такой слой есть. 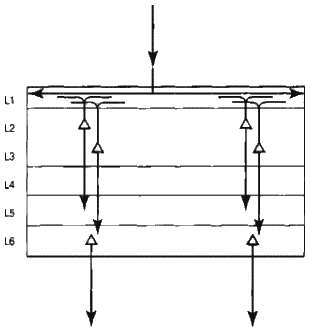 Рис. 6.8. Нисходящий информационный поток И еще немного сведений из анатомии: аксоны, покидающие слой 6, окутаны оболочкой из белого жирового вещества, которое называется миелином. Это белое вещество выполняет ту же роль, что и изоляция на электрических проводах, – препятствует смешиванию сигналов и повышает скорость их передвижения до 320 километров в час. Аксоны избавляются от белого защитного вещества, попадая в новую колонку нейронов слоя 6 в коре головного мозга. И наконец, есть еще один непрямой способ связи зон коры головного мозга. Но прежде я хотел бы вам снова напомнить об автоассоциативном свойстве системы памяти (о котором шла речь в главе 2). Как вы, вероятно, помните, автоассоциативные запоминания служат для сохранения последовательностей сигналов. Когда выходной сигнал группы искусственных нейронов передается назад с целью формирования входного потока для всех нейронов (и при этом обратная связь дополняется задержкой), то таким образом сигналы учатся последовательно двигаться друг за другом. По моему мнению, кора головного мозга использует подобный механизм для сохранения последовательностей, но в нем присутствуют еще некоторые особенности. Вместо того чтобы формировать автоассоциативные последовательности из искусственных нейронов, она формирует их из колонок коры головного мозга. Исходящие данные из всех колонок передаются назад, в слой 1. Выходит, слой 1 располагает свежей информацией о том, какие колонки зоны коры головного мозга только что были активными. Рассмотрим элементы, изображенные на рис. 6.9. Ученым давно известно, что нейроны особо крупных размеров слоя 5 моторной коры (зона M1) непосредственным образом связаны с мышцами и моторными зонами спинного мозга – они обеспечивают управление вашими мышцами и передвижение. Каждый раз, когда вы говорите, печатаете, выполняете любые самые сложные действия, эти клетки посылают высококоординированные импульсы, заставляя сокращаться ваши мышцы. Совсем недавно ученые открыли, что крупные нейроны слоя 5, возможно, выполняют какие-то функции и в других частях коры головного мозга, а не только в моторных зонах. Например, крупные нейроны слоя 5 зрительной зоны имеют проекции в зоне, отвечающей за движения глаз. Сенсорные зрительные зоны V2 и V4 не только занимаются обработкой зрительных входных сигналов, но также помогают предопределять движение глаз, а следовательно, управляют тем, что вы видите. Крупные нейроны слоя 5 присутствуют в каждой зоне коры головного мозга, что наводит на мысль об их немаловажной роли во всех типах движений. Аксоны этих нейронов не только участвуют в формировании поведения, они разветвляются надвое. Одно ответвление идет к таламусу, изображенному на рис. 6.9 в виде овальной фигуры. Таламус человека находится в центре головного мозга, под корой, сверху «старого мозга», и окружен белым веществом. Размером и формой он похож на два маленьких птичьих яйца. Все зоны коры головного мозга направляют многочисленные аксоны в таламус, от него тоже исходят аксоны в обратном направлении. Науке известно достаточно об этих связях, но сам таламус очень сложен по своей структуре, и его роль до сих пор мало изучена. Таламус необходим для того, чтобы вести нормальный образ жизни: его повреждение приводит к устойчивому «растительному» состоянию. 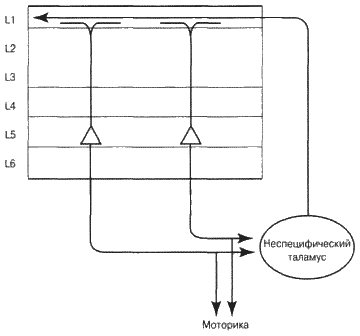 Рис. 6.9. Как посредством таламуса сообщаются текущее состояние и текущее моторное поведение Существуют несколько путей от таламуса к коре головного мозга, но сейчас лишь один из них представляет для нас интерес. Он начинается с крупных нейронов слоя 5, проецирующихся на группу клеток таламуса, считающихся функционально неспецифическими. Аксоны этих неспецифических клеток идут назад, в слой 1 множества различных зон коры головного мозга. Например, нервные клетки слоя 5 зон V2 и V4 посылают аксоны к таламусу, а таламус, в свою очередь, отправляет информацию назад к слою 1 зон V2 и V4. Аналогичный процесс происходит и в других зонах коры головного мозга. Клетки слоя 5 множества корковых зон посылают сигналы в таламус, который, в свою очередь, отсылает назад информацию в слой 1 этих же самых и связанных с ними зон. Я считаю, что такой круговорот полностью дублирует обратную связь с задержкой, которая позволяет автоассоциативной памяти усваивать последовательности сигналов. Только что я упомянул о двух источниках входной информации, поступающей в слой 1. Высшие зоны коры головного мозга возбуждают активность в слое 1 более низких зон коры. Активные колонки в пределах одной зоны возбуждают активность в пределах слоя 1 той же зоны через таламус. Эти входящие в слой 1 сигналы можно представить как название песни – считать словами песни (входящие сверху сигналы) и знание, в каком месте песни мы сейчас находимся (задержка в деятельности активных колонок в пределах одной зоны). Таким образом, слой 1 содержит большое количество информации, необходимой нам для прогнозирования времени активизации колонки: название последовательности и наше местонахождение в ней. Используя эти два сигнала слоя 1, зона коры головного мозга в состоянии обучаться и запоминать огромное количество последовательностей сигналов. Как работает корковая зона: подробностиНе забывая о наших трех схемах (входные сигналы, поступающие вверх по иерархии; разветвляющиеся сигналы, спускающиеся вниз по иерархии; обратная связь с задержкой в таламусе), приступим к рассмотрению того, как зона коры головного мозга выполняет свои функции. Нам необходимо выяснить следующее. Как зона мозга классифицирует входящие сигналы (вспомните сортировку цветных листов бумаги по ведрам). Как она усваивает последовательности сигналов (вспомните последовательности интервалов в песне и восприятие лиц посредством комбинаций типа «глаз-нос-глаз»). Как корковая зона формирует постоянные паттерны, или «имена» последовательностей. Как она составляет специфические прогнозы (встреча поезда в определенное время или прогнозирование следующего звука мелодии). Допустим, что колонки одной зоны – это те же ведра, которые мы использовали для воображаемой сортировки получаемых цветных листов бумаги. Каждая колонка помечена точно так же, как и ведро. К клеткам слоя 4 в каждой колонке протянуты волокна ввода из нескольких зон, расположенных ниже по иерархии. Всякий раз, получая правильную комбинацию входных сигналов, они выдают импульсный разряд. Возбуждаясь, клетка слоя 4 «голосует» за то, что полученные сигналы соответствуют своему ярлыку. Точно так же, как при сортировке листов бумаги, входные сигналы могут быть неоднозначными (например несколько колонок могут подходить для единственного входного сигнала). Мы хотим, чтобы зона коры нашего головного мозга сделала выбор: лист бумаги может считаться либо красным, либо оранжевым, но ни в коем случае не тем и другим сразу. Колонка, которой входной сигнал соответствует более всего, должна предотвратить активизацию остальных колонок. Этим занимаются так называемые тормозные нейроны. Они предотвращают активизацию соседних нервных клеток, эффективно заботясь о том, чтобы был только один «победитель». Эти тормозные нейроны влияют исключительно на зону вокруг «своей» колонки. Другими словами, несмотря на наличие механизма подавления, существует возможность одновременной активизации нескольких колонок. (В мозге ничто не бывает представлено одним единственным нейроном или единственной колонкой.) Для простоты понимания можем представить, что зона выбирает одну и только одну колонку-«победителя», однако на будущее запомним, что на самом деле одновременно активизируются множество колонок. Процесс, используемый корой головного мозга для классификации входных сигналов, сложен и плохо изучен. Я не собираюсь заводить вас в дебри. Вместо этого я предлагаю принять предположение, что зона коры головного мозга отсортировала входной сигнал как активизацию в наборе колонок. Тогда мы сможем сосредоточиться на формировании последовательностей и присвоении им названий. Каким образом зона коры головного мозга сохраняет последовательность отсортированных сигналов? Я уже излагал свои соображения по этому поводу и сейчас хотел бы углубиться в подробности. Представьте себе, что вы – колонка нервных клеток, а сигнал, поступивший из зоны низшего порядка, заставил активизироваться одну из ваших клеток слоя 4. Вслед за этим возбуждаются клетки в слоях 2 и 3, потом – в слое 5, а затем – в 6-м. Постепенно активизируется вся колонка нейронов. У каждой из клеток слоев 2, 3 и 5 есть тысячи синапсов в слое 1. Если какие-то из этих синапсов активны во время разряда нервных клеток слоев 2, 3 и 5, они усиливаются. Если такое происходит часто, то синапсы слоя 1 становятся достаточно сильными для того, чтобы вызвать вспышки клеток в слоях 2, 3 и 5 даже тогда, когда клетки слоя 4 молчат. Таким образом, некоторые части колонки могут активизироваться, даже если они не получили входного сигнала от более низкой по иерархии зоны коры головного мозга. Именно так клетки слоев 2, 3 и 5 учатся предвосхищать собственную активизацию на основе сигнала, поступающего из слоя 1. «До обучения» колонка может активизироваться исключительно через клетки слоя 4. «После обучения» колонка может частично активизироваться через память. Когда колонка активизируется через синапсы слоя 1, то входящие в нее нейроны предвидят, что произойдет активизация снизу, – это и есть прогноз. Если бы колонка могла разговаривать, она бы сказала: «Когда я активизировалась в прошлом, то как раз этот набор моих синапсов слоя 1 был активен. Как только я увижу такой набор синапсов, сразу же начну генерировать импульс в предвкушении сигнала». Вспомните, что половина входных сигналов слоя 1 происходит из слоя 5 клеток соседних колонок и других зон коры головного мозга. Это информация о том, что случилось мгновение тому назад. Она говорит нам, какие колонки были активными до того, как активизировалась наша колонка. Она говорит нам, каким был последний интервал мелодии, что я видел последним, что я чувствовал последним, какую фонему устной речи я услышал последней и так далее. Если порядок возникновения указанных сигналов является последовательным во времени, то колонки усваивают последовательность. Они будут возбуждаться одна за другой в определенной последовательности. Вторая половина сигналов поступает в слой 1 из клеток слоя 6 зон, расположенных выше в иерархии. Эта информация является более стабильной и представляет «имя» последовательности, с которой вы непосредственно сталкиваетесь в данный момент времени. К примеру, если колонки представить как музыкальные фразы, то название мелодии будет «именем» последовательности. Если фонемы будут колонками, то «именем» будет слово, которое вы слышите. Если колонками будут произносимые вами слова, «именем» будет речь, которую вы произносите. Следовательно, в слое 1 содержится и информация об «имени» последовательности, и о последнем сигнале в ней. Благодаря этому одна колонка может использоваться многими последовательностями без возникновения недоразумений. Колонки «учатся» активизироваться в нужном контексте и в правильном порядке. Я должен подчеркнуть, что синапсы слоя 1 не являются единственными синапсами, участвующими в процессе обучения тому, когда должна активизироваться колонка. Как я уже упоминал ранее, клетки получают и отсылают сигналы ко многим соседним колонкам. Ведь более 90% всех синапсов – соединения с клетками за пределами колонки, и большинство из них расположены не в слое 1. Например, у клеток слоев 2, 3 и 5 есть тысячи синапсов в слое 1, но, кроме того, они имеют и тысячи синапсов в собственных слоях. Общий принцип состоит в том, что нейроны стремятся получать любую информацию, которая поможет им спрогнозировать возникновение импульса снизу. Как правило, активность соседних колонок отличается высоким уровнем корреляции, поэтому существует много связей между соседними колонками. Например, если в поле вашего зрения находится прямая линия, это активизирует колонки одну за другой. Но достаточно часто для прогнозирования активности колонки нужна более обширная информация. Именно тогда вступают в игру синапсы слоя 1. Нервным клеткам или колонкам нейронов не известно, что означает тот или иной синапс, но посредством синапсов они прогнозируют, когда нужно активизироваться. А сейчас давайте рассмотрим, как зона коры головного мозга формирует имя выученной последовательности. Опять представьте себя зоной коры головного мозга. Активность ваших колонок нейронов меняется при каждом новом входном сигнале. Вы успешно усвоили последовательность активизации ваших колонок, т. е. некоторые клетки ваших колонок активизируются до того, как поступит входной сигнал из более низких зон. Какую информацию вы посылаете в более высокие зоны коры головного мозга? Как мы знаем, клетки слоев 2 и 3 посылают свои аксоны в следующую вверх по иерархии область. Активность этих клеток является входным сигналом для более высоких областей. Вот тут и возникает проблема. Чтобы иерархия работала слаженно, вам нужно транслировать постоянный сигнал на протяжении всей выученной последовательности. Вы должны транслировать «имя» последовательности, а не ее детали. До того как вы «выучите» последовательность, вы можете передавать отдельные ее элементы, но, после того как вы ее усвоили и можете прогнозировать, какие колонки будут активизироваться, вы должны передавать постоянный сигнал. Однако я еще не рассказывал, как это происходит. Итак, пока что вы передаете каждый переменный сигнал независимо то того, можете вы его спрогнозировать или нет. Когда активизируется вся колонка, клетки слоев 2 и 3 посылают новый сигнал вверх по иерархии. Коре головного мозга нужен какой-то способ, позволяющий поддерживать постоянный сигнал на протяжении всей выученной последовательности. Нам нужен способ отключать сигналы, исходящие из слоев 2 и 3, когда колонка прогнозирует свою активность, или же, наоборот, активизировать эти клетки, когда колонка не может спрогнозировать свою активность. Только так можно создать постоянный сигнал имени. 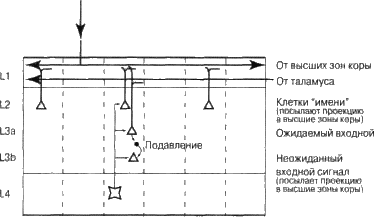 Рис. 6.10. Формирование постоянного «имени» для усвоенной последовательности Науке очень мало известно о том, как именно кора головного мозга осуществляет подобную операцию. По моему мнению, существует несколько потенциально возможных методов. Я опишу тот, к которому больше всего склоняюсь в последнее время. (Не забывайте, что для нас сейчас наиболее важными являются концептуальные рамки, а не конкретный метод.) Ключевым моментом является создание постоянного сигнала «имени». Я хочу лишь показать, что существуют вполне правдоподобные механизмы для осуществления такого процесса. Представьте, что вы – колонка (см. рис. 6.10). Постараемся разобраться, как вы учитесь посылать постоянный сигнал в вышестоящую в иерархии область, когда вам удается спрогнозировать свою активность, и переменный сигнал в противном случае. Сначала предположим, что в слои 2 и 3 входят несколько классов клеток. (Впридачу к нескольким типам тормозных нейронов многие анатомы различают несколько типов в слоях За и 3б, т. е. наше предположение вполне оправдано.) Теперь предположим, что один класс клеток – клетки слоя 2 – учится быть активным на протяжении усвоенных последовательностей. Как группа этих клеток представляет «имя» последовательности? Она посылает постоянный сигнал в высшие зоны коры головного мозга до тех пор, пока их зона может предвидеть, какие колонки будут активизироваться дальше. Если ваша зона усвоила последовательность из трех различных сигналов, тогда клетки слоя 2 всех колонок, представляющих эти три сигнала, будут оставаться активными до тех пор, пока мы остаемся в пределах нашей последовательности. Они и являются «именем» последовательности. Теперь предположим, что существует другой класс клеток – клетки слоя 3б, которые не активизируются, когда наша колонка успешно прогнозирует поступающие сигналы, но генерируют разряд в случае, если этого не происходит. Клетка слоя 3б олицетворяет' неожиданный сигнал. Она генерирует импульс при неожиданной активизации своей колонки. Она будет поступать так всякий раз, когда колонка, еще не «прошедшая» процесс обучения, активизировалась. По мере того как колонка учится прогнозировать свою активность, клетки слоя 36 «успокаиваются». Вместе взятые, клетки слоя 2 и слоя 3б удовлетворяют нашим требованиям. До того как было пройдено «обучение», и те и другие клетки затухают-вспыхивают в колонке, но по окончании «учебного курса» клетка слоя 2 остается активной постоянно, а клетка слоя 36 пребывает в состоянии покоя. Как клетки учатся этому? Сначала давайте рассмотрим, как «отключить» клетки слоя 3б, когда колонка успешно прогнозирует свою активность. Допустим, существует клетка За, расположенная над клеткой 36, она имеет дендриты в слое 1. Ее единственная задача состоит в том, чтобы воспрепятствовать активизации клетки слоя 36, если она замечает в слое 1 уже известный паттерн. Когда клетка слоя За видит выученный сигнал в слое 1, она сразу же активизирует тормозной нейрон, который, в свою очередь, препятствует вспышке клетки слоя 36. Этого достаточно, чтобы удержать клетку слоя 3б от вспышки, когда колонка правильно прогнозирует свою активность. А теперь рассмотрим более сложную задачу: как клетка слоя 2 остается активной на протяжении всей известной последовательности сигналов. Данная задача более сложна, потому что при этом различные клетки слоя 2 из многих колонок должны одновременно оставаться активными, даже если отдельные колонки в текущий момент таковыми не являются. По моему мнению, это может происходить следующим образом. Клетки слоя 2 учатся активизироваться исключительно высшими областями иерархии коры головного мозга. Они формируют синапсы преимущественно с аксонами из клеток слоя 6 областей, расположенных на уровень выше. Таким образом, клетки слоя 2 представляют постоянный сигнал «имени» из высшей области. Когда высшая зона посылает сигнал вниз к слою 1 расположенной ниже области, набор клеток слоя 2 нижней зоны активизируется, представляя все колонки, являющиеся членами последовательности. Клетки слоя 2 тоже имеют проекции в вышестоящей области и представляют собой частично стабильную группу клеток. (Вряд ли они постоянно остаются активными. Скорее всего, они вспыхивают синхронно, наподобие ритма.) Верхняя зона как бы посылает название мелодии в слой 1, расположенный ниже. Это заставляет вспыхивать набор клеток слоя 2, каждую клетку в своей колонке, и они будут активизироваться, когда вы слышите мелодию. Все эти механизмы вместе взятые позволяют коре головного мозга усваивать последовательности, составлять прогнозы, формировать постоянные репрезентации («имена») для последовательностей входящих сигналов. Это основные операции, необходимые для формирования инвариантных репрезентаций. Как мы формируем прогнозы о событиях, с которыми ранее не сталкивались? Как мы выбираем одну из десятков возможных интерпретаций входных сигналов? Как зона коры головного мозга формирует частные прогнозы на основе инвариантных схем? Мы уже рассматривали примеры прогнозирования следующего звука мелодии, когда в вашей памяти есть только интервал между отдельными нотами, историю со встречей поезда и воспроизведение Геттисбергской речи. Во всех перечисленных случаях единственным путем решения проблемы было использование последней полученной информации для того, чтобы конвертировать инвариантный прогноз в частное предсказание. Перефразировав ту же мысль в терминах коры головного мозга, можем сказать, что нужно сочетать поток прямой информации (поступающие фактические сигналы) с потоком обратной информации (прогноз в инвариантной форме). Приведу простой пример того, как это, по моему представлению, происходит. Допустим, кора вашего головного мозга знает, что должна воспринять музыкальный интервал квинту. Колонки вашей зоны реагируют на отдельные интервалы, такие как «до-ми», «до-соль», «ре-ля» и так далее. Вам нужно решить, какие из колонок будут активными. Если зона выше говорит вам, что следует ожидать квинту, это активизирует клетки слоя 2 во всех колонках, представляющих квинты, то есть «до-соль», «ре-ля», «ми-си». Клетки слоя 2 из колонок, которые представляют другие интервалы, активизироваться не будут. Теперь вам нужно из колонок, представляющих квинты, выбрать только одну. Входными сигналами для вашей зоны будут отдельные ноты. Если «ре» была последней нотой, которую вы слышали, тогда все колонки, представляющие интервалы, в которых присутствует нота «ре», т. е. «ре-ми» и «ре-си», получают часть входного сигнала. Теперь в слое 2 активизировались все колонки, представляющие квинты, а в слое 4 мы получили часть сигнала во все колонки, представляющие интервалы, в которые входит нота «ре». Пересечение этих двух множеств и будет ответом на наш вопрос. Это будет колонка, представляющая интервал «ре-ля» (см. рис. 6.11). Как кора головного мозга определяет пересечение? Мы раньше упоминали о том, что аксоны клеток слоев 2 и 3, выходя из коры головного мозга, формируют синапсы в слое 5. Аксоны, направляющиеся к слою 4 из более низких зон коры головного мозга, создают синапсы в слое 6. Пересечение этих двух синапсов (снизу вверх и сверху вниз) дает нам как раз то, что нужно. Клетки слоя 6, получающие эти два сигнала возбуждаются. Клетка слоя 6 «представляет мнение» зоны коры головного мозга о том, что происходит, т. е. специфический прогноз. Если бы клетка слоя 6 могла разговаривать, она бы сказала: «Я являюсь частью колонки, представляющей нечто. Конкретно в моем случае колонка представляет интервал „ре-ля“. У остальных колонок другие значения. Я представляю свою зону коры головного мозга. Когда я становлюсь активной, это означает, что, по нашему мнению, сейчас звучит интервал „ре-ля“, или же он прозвучит в ближайшем будущем. Возможно, я активизировалась, потому что сигнал снизу вверх, поступивший от ушей, заставил клетку слоя 4 моей колонки возбудить всю колонку. Или же моя активизация может означать, что мы узнали мелодию и прогнозируем следующий интервал. В любом случае моя задача состоит в том, чтобы довести до ведома низших зон коры головного мозга, что, по нашему мнению, происходит. Я представляю наше видение внешнего мира – независимо от того, является он настоящим или выдуманным». Попробую описать указанный процесс с помощью визуальной картинки. Представьте себе два листа бумаги, усеянные множеством мелких дырочек. Дырочки на одном листе представляют колонки, у которых есть активные клетки слоев 2 и 3 (наш инвариантный прогноз). Дырочки на другом листе бумаги представляют колонки с частичным сигналом снизу. Если мы наложим листы бумаги друг на друга, то некоторые из дырочек совпадут, а некоторые – нет. Дырочки, которые совпали, представляют колонки, которые, как мы полагаем, должны активизироваться. 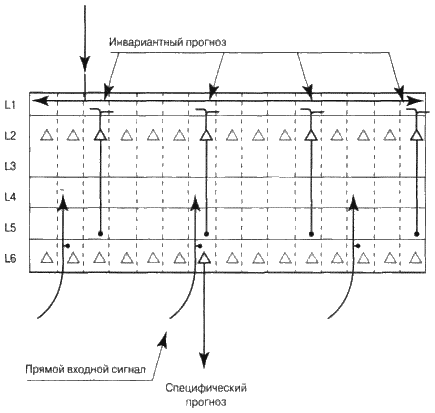 Рис. 6.11. Как зона коры головного мозга формирует специфические прогнозы, используя инвариантные репрезентации Описанный механизм не только создает определенные прогнозы, он также разрешает неоднозначность входных сенсорных сигналов. Как мы помним из примеров с цветными листами бумаги и искаженным изображением, очень часто входные сигналы, полученные зоной коры головного мозга, являются неоднозначными. Механизм сочетания информации, поступающей снизу вверх и сверху вниз, помогает выбрать один из двух или более вариантов интерпретации. Как только вы приняли решение, каков будет выбор, вы сразу же транслируете этот выбор в область, расположенную ниже по иерархии. Каждое мгновение нашей жизни каждая зона коры головного мозга сравнивает набор ожидаемых колонок, возбужденных сверху, с набором колонок, которые уже были возбуждены снизу. Там, где эти два набора пересекаются, создается наше восприятие. Если бы поступали совершенные сигналы снизу и у нас были бы совершенные прогнозы, тогда набор возбужденных колонок всегда совпадал бы с набором ожидаемых колонок. Но очень часто этого не происходит. Метод сочетания частичного прогноза и частичного входного сигнала позволяет определиться с неоднозначным входным сигналом, он позволяет заполнить пробелы в информации и остановить выбор на одном из альтернативных вариантов. Именно так мы комбинируем музыкальный интервал с последней услышанной нотой, чтобы спрогнозировать следующую ноту мелодии. Именно так мы определяемся, изображено на картинке два лица или ваза. Именно так мы разветвляем моторный поток на написание или произнесение вслух Геттисбергской речи. И наконец, вдобавок к отправлению информации в более низкие зоны коры головного мозга, клетки слоя 6 могут посылать выходные сигналы назад, к клеткам слоя 4 той же колонки. Когда они это делают, наши прогнозы становятся входным сигналом. Именно этим мы занимаемся, размышляя или мечтая. Это позволяет нам видеть последовательности наших собственных прогнозов. Мы занимаемся этим, планируя свое будущее, вспоминая выступления или переживая о предстоящих событиях. Стефен Гросберг, длительное время занимавшийся моделированием деятельности коры головного мозга, называет это свернутой обратной связью, я же предпочитаю термин воображение. И наконец, последний вопрос, который нам нужно затронуть, прежде чем перейти к следующему разделу. Я неоднократно подчеркивал, что чаще всего то, что мы видим, слышим, чувствуем, во многом зависит от наших собственных действий. То, что мы видим, зависит от того, на чем наши глаза остановились после очередной саккады, и от угла поворота головы. То, что мы ощущаем, зависит от того, как мы передвигаем пальцы и конечности. То, что мы слышим, иногда зависит от того, что мы делаем и говорим. Значит, чтобы спрогнозировать, что мы почувствуем дальше, мы должны знать, что мы сейчас делаем. Моторное поведение и сенсорное восприятие тесно взаимосвязаны. Как нам удается составлять прогнозы, если то, что мы чувствуем, во многом является результатом наших собственных действий? К счастью, у данной проблемы существует неожиданное и простое решение, правда, многие его детали не очень понятны. Неожиданное открытие: восприятие и поведение – это почти одно и то же. Как я уже упоминал раньше, большинство зон коры головного мозга (если не все) принимают участие в осуществлении движения. Клеткам слоя 5, имеющим проекции в таламусе, а затем в слое 1, тоже свойственна моторная функция, поскольку у них одновременно есть проекции в моторных зонах «старого» мозга. Таким образом, знание о том, что только что произошло (как в сенсорном, так и в моторном плане), присутствует в слое 1. Вторая неожиданность (являющаяся следствием первой): моторное поведение должно быть представлено в иерархии инвариантных репрезентаций. Вы генерируете движения, необходимые для осуществления определенного действия, не размышляя о нем во всех деталях. Когда моторная команда опускается вниз по иерархии, она трансформируется в сложные и подробные последовательности, необходимые для осуществления той деятельности, которая была запланирована. Это происходит как в сенсорной, так и в моторной зонах коры головного мозга, что размывает грань между ними. Если зона IT зрительной коры головного мозга воспринимает репрезентацию «нос», то простое действие переключения на репрезентацию «глаза» будет вызывать саккаду, необходимую для оправдания прогноза. Саккада, необходимая для того, чтобы перейти от зрительного восприятия носа к зрительному восприятию глаза, меняется в зависимости от расположения лица. Если лицо расположено поблизости, это требует большей саккады, и наоборот, более отдаленное лицо требует меньшей саккады. Наклоненное лицо требует саккады под углом, который отличается от используемого для лица без наклона. То, какой должна быть саккада, определяется, пока прогноз о том, что следующим мы увидим глаз, спускается к зоне V1. Чем дальше опустился прогноз, тем более точной становится саккада. Заканчивается все тем, что центральные ямки ваших глаз останавливается точно на объекте или же очень близко к нему. Давайте рассмотрим еще один пример. Для того чтобы я физически переместился из гостиной на кухню, все, что нужно моему мозгу, – это мысленно переключиться с инвариантной репрезентации гостиной на инвариантную репрезентацию кухни. Данное переключение вызывает сложное разворачивание последовательностей. Процесс генерирования последовательности прогнозов того, что я увижу, услышу, почувствую, пока буду переходить из гостиной на кухню, тоже генерирует последовательность моторных команд, заставляющих меня перейти из гостиной на кухню и перемещать мой взгляд, пока я двигаюсь. Прогнозирование и моторное поведение идут рука об руку, в то время как потоки сигналов одновременно двигаются вверх и вниз по иерархии коры головного мозга. Каким бы странным это ни казалось, но, когда в дело вовлечено ваше поведение, ваши прогнозы не только предшествуют ощущению – они также его предопределяют. Прогнозирование перехода к следующему сигналу последовательности вызывает каскадное прогнозирование того, что вы должны пережить дальше. Когда каскадный прогноз разворачивается, он генерирует моторные команды, необходимые для осуществления прогноза. Воображение, прогнозирование и действие являются частями одного и того же разворачивания последовательностей в иерархии коры головного мозга. Действие в воображении, параллельное разворачивание сенсорного и моторного поведения являются основой поведения, ориентированного на результат. Целенаправленное поведение, этот священный Грааль робототехники, изначально встроено в ткань коры головного мозга. Конечно, мы можем отключить наше моторное поведение. Я могу представить себе, что вижу что-то, хотя на самом деле этого не вижу. Я могу представить себе, как иду на кухню, хотя на самом деле этого не делаю. Но действие всегда начинается в нашем воображении. Поток вниз, поток вверхДавайте на минуту остановимся и поразмышляем над тем, как информация перемещается вниз и вверх в иерархии коры головного мозга. Когда вы передвигаетесь, потоки непрерывно меняющейся входной информации устремляются в нижние зоны коры головного мозга. Каждая зона пытается интерпретировать поток входящей информации как часть известной последовательности сигналов. Колонки пытаются предугадать свою активность. Если им это удается, они передают постоянный сигнал – название последовательности – в область, расположенную выше в иерархии. Зона коры головного мозга как бы говорит: «Я слышу песню, и вот вам ее название. Я сама разберусь с отдельными элементами». Но что будет, если поступит неожиданный сигнал – нота, которой вы не ожидали? Что будет, если мы увидим что-то такое, чего на лице быть не должно? Неожиданный сигнал автоматически будет передан в область, стоящую выше в иерархии. Это случается, когда «вспыхивают» клетки слоя 3б, которые не были частью ожидаемой последовательности. Возможно, более высокая зона сможет понять сигнал как часть ее собственной последовательности. Возможно, она скажет: «Я вижу, что тут появилась новая нота. Вполне возможно, это первая нота следующей песни из альбома. Мне так кажется, поэтому я прогнозирую, что мы перешли к следующей песне из альбома. Низшая область, вот тебе название песни, которую, как мне кажется, ты сейчас слышишь». Но если не произойдет такого распознания, сигнал будет подниматься вверх по иерархии коры головного мозга, пока какая-нибудь из вышестоящих зон не идентифицирует сигнал как часть нормальной последовательности событий. Чем выше по иерархии поднимается сигнал, тем больше разных зон вовлекается в его обработку. Когда же наконец какая-то зона в верхней части иерархии считает, что она в состоянии обработать неожиданное событие, тогда формируется новый прогноз. Новый сигнал продвигается вниз по иерархии так далеко, как ему удается. Если новый прогноз оказался неверным, ошибка будет выявлена, и сигнал снова начнет подниматься вверх по иерархии до тех пор, пока не найдется область, которая интерпретирует его как часть своей непосредственной активной последовательности. Отсюда вывод: сигналы передвигаются вверх по иерархии, а прогнозы – вниз. В идеале, в хорошо известном и прогнозируемом мире, эти встречные потоки сигналов быстротечны и циркулируют преимущественно в нижних областях коры головного мозга. Мозг как можно быстрее пытается найти ту часть своей модели мира, которая бы соответствовала неожиданному входному сигналу. Только после этого он может понять входной сигнал и знать, чего ожидать дальше. Когда я хожу по знакомой комнате в своем доме, лишь несколько ошибок поднимутся к верхним областям коры головного мозга. Хорошо усвоенные последовательности сигналов в моем доме обрабатываются на уровне нижних зон зрительной, соматосенсорной и моторной иерархии. Я знаю комнату настолько хорошо, что могу передвигаться по ней даже в кромешной тьме. Благодаря тому, что я хорошо знаю обстановку вокруг меня, большая часть моего неокортекса свободна и может размышлять о мозге и написании книг. В то же время если бы я находился в незнакомой мне комнате, особенно в такой, подобной которой я никогда не встречал, то мне не только пришлось бы постоянно присматриваться, куда идти. Неожиданные сигналы мощным потоком ринулись бы к верхним областям коры головного мозга. Чем больше мой сенсорный опыт не соответствует заученным последовательностям, тем больше ошибок будет возникать. В такой непривычной ситуации я не смогу думать о мозге, поскольку большая часть коры головного мозга будет занята проблемами ориентирования в комнате. С подобными ощущениями сталкиваются люди, сошедшие с трапа самолета в незнакомой стране. Дороги похожи на те, к которым вы привыкли, автомобили придерживаются левой стороны, язык, на котором все общаются, вам непонятен, а поиск туалета может стать сверхзадачей для всей коры головного мозга. Даже не пытайтесь готовиться к публичному выступлению, разгуливая по незнакомой стране. Ощущение внезапного понимания (момент озарения) можно объяснить в рамках модели. Представьте, что вы смотрите на неоднозначную картинку. Пятна чернил и какие-то линии ничего вам не напоминают. Полнейшая бессмыслица. Когда мозг не может найти воспоминания, соответствующего полученному сигналу, человек пребывает в замешательстве. Ваши глаза блуждают по картинке. Входные сигналы поднялись на самый верх иерархии. Высшие зоны выдвигают множество самых разных гипотез, но, как только прогнозы начинают опускаться вниз по иерархии, каждая из гипотез конфликтует с входными сигналами, и мозг вынужден начинать все сначала. На протяжении всего состояния замешательства ваш мозг полностью поглощен тем, что пытается понять картинку. Наконец он формирует прогноз, который оказывается правильным. Когда это происходит, прогноз, сформированный в верхней зоне коры головного мозга, стремительно спускается вниз. Менее чем за секунду каждая зона получает последовательность, соответствующую полученным данным. Ни одна ошибка больше не транслируется к верхним зонам иерархии. Вы понимаете картинку и вместо точек и пятен видите далматинца (рис. 6.12). На что способна обратная связь?Десятки лет нам было известно, что связи в коре головного мозга являются взаимными. Если у зоны А есть проекция в зоне Б, то у зоны Б есть проекция в зоне А. Часто аксонов обратной связи оказывается больше, чем волокон прямой связи. Несмотря на то что все признают существование такого феномена, считается, что обратная связь играет второстепенную, или «модуляторную», роль в работе мозга. Мысль о том, что сигнал обратной связи не может постоянно и точно активизировать различные группы клеток слоя 2, не имеет большой поддержки в рядах нейробиологов. Почему так? Частично потому, что, как я уже упоминал, если вы не считаете функцию прогнозирования центральной функцией головного мозга, то не возникает особой необходимости обращать внимание на обратную связь. Если вы предполагаете, что информация движется прямолинейно, к моторной системе, зачем вам эта обратная связь? Еще одна причина игнорирования обратной связи – то, что сигнал обратной связи широко распространяется в слое 1. Интуитивно мы ожидаем, что сигнал, рассеянный в большой зоне, будет оказывать очень слабое влияние на большое количество нейронов. В самом деле, у мозга есть несколько таких модуляторных сигналов, которые не оказывают влияния на отдельные нейроны, зато меняют общие характеристики (например, возбудимость).  Рис. 6.12. Вы видите далматинца? И последняя причина игнорирования обратной связи обусловлена тем, как многие нейробиологи представляют себе работу нейронов. У типичных нейронов имеются тысячи или даже десятки тысяч синапсов. Одни из них расположены ближе к телу клетки, другие – дальше. Синапсы, расположенные ближе к телу клетки, оказывают сильное влияние на ее импульсацию. Дюжина активных синапсов, расположенных у тела клетки, может заставить ее генерировать потенциал действия – электрический разряд. В этом не ничего нового. Тем не менее большинство синапсов находятся вдали от тела клетки. Они разбросаны по разветвленной структуре дендритов клетки. Поскольку эти синапсы расположены далеко от тела клетки, ученые склонны полагать, что импульс, пришедший на один из этих синапсов, будет очень слабо либо вообще не будет влиять на активность клетки. Импульс с отдаленного синапса рассеется по дороге, не достигнув тела клетки. Как правило, информация, поднимающаяся вверх по иерархии коры головного мозга, передается по синапсам, расположенным близко к телу клетки. В этом случае сведения с большей вероятностью будут успешно передаваться от зоны к зоне. Как правило, обратная информация, поступающая вниз по иерархии коры головного мозга, передается по синапсам, расположенным дальше от тела клетки. Клетки слоев 2, 3 и 5 посылают дендриты в слой 1 и формируют там много синапсов. Слой 1 представляет собой совокупность синапсов, но все они значительно отдалены от тел клеток слоев 2, 3 и 5. Более того, каждая отдельная клетка слоя 2 формирует чрезвычайно мало синапсов с каждым отдельным волокном обратной связи (если формирует их вообще). Следовательно, некоторые ученые могут возразить по поводу того, что непродолжительный сигнал в слое 1 может вызвать упорядоченное возбуждение группы клеток слоев 2, 3 и 5, однако моя теория базируется именно на этом предположении. Дилемма разрешается благодаря тому, что фактическое поведение нейронов отличается от поведения, приписываемого им классической моделью. В последние годы многие ученые придерживаются мнения, что синапсы на отдаленных тонких дендритах могут играть самую непосредственную роль в возбуждении клеток. В указанных моделях эти дальние синапсы функционируют не так, как синапсы на более толстых дендритах, расположенных вблизи тела клетки. Например, если два синапса на тонком дендрите расположены очень близко друг к другу, они будут действовать как «детектор совпадения». Другими словами, если оба синапса получили входящий импульс близко во времени, то они могут оказать большое влияние на клетку, хотя и расположены далеко от ее тела. Они могут заставить тело клетки генерировать разряд. Поведение дендритов нейронов до сих пор остается загадкой для науки, поэтому я не могу остановиться на этом вопросе более подробно. Важным моментом является то, что модель коры головного мозга «Память-предсказание» требует, чтобы синапсы, отдаленные от тела клетки, могли идентифицировать специфические сигналы. Не слишком прозорливым, почти глупым было заявление, что большинство из тысяч синапсов нейрона играют только модуляторную роль. Обширная обратная связь и огромное количество синапсов не могли возникнуть без веской причины. Используя наши знания, мы можем сказать, что типичный нейрон обладает способностью выучить сотни точных совпадений благодаря волокнам обратной связи, когда они создают синапсы на тонких дендритах. Это, в свою очередь, означает, что колонка коры головного мозга очень гибка в отношении того, какие сигналы обратной связи могут вызвать ее активность. Таким образом, каждое отдельное свойство можно точно ассоциировать с тысячами различных объектов и последовательностей. Моя модель требует того, чтобы обратная связь была быстрой и точной. Клетки должны возбуждаться, когда они обнаруживают любое количество точных совпадений на отдаленных дендритах. Новая нейронная модель основывается именно на этом. Как происходит процесс обученияУ всех нейронов из всех слоев коры головного мозга есть синапсы, большинство из которых может модифицироваться под влиянием опыта. Можно с уверенностью сказать, что процесс обучения и запоминания происходит во всех слоях, во всех колонках и во всех зонах коры головного мозга. Я упоминал ранее правило «обучения Хебба», названное так в честь Дональда Хебба. Сущность этого правила очень проста: когда два нейрона активизируются одновременно, синаптические связи между ними усиливаются. (Его легко запомнить как фразу «Возбуждены вместе – связаны вместе».) Сейчас мы знаем, что Хебб был прав. Конечно, ничего в природе не бывает настолько простым, свойства настоящего мозга сложнее. Наша нервная система работает на основе многих вариаций «обучения Хебба». Некоторые синапсы изменяют силу связи в ответ на незначительные отклонения координации нейронных сигналов. Некоторые синаптические изменения оказываются краткосрочными, некоторые – долгосрочными. Хебб очертил основные рамки, а не создал конечную теорию процесса обучения, причем эти рамки оказались очень и очень полезными. Принципы «обучения Хебба» объясняют большинство проявлений поведения коры головного мозга, о которых мы говорили в данной главе. Помните, еще в 1970-х годах было установлено, что автоассоциативная память при помощи алгоритма «обучения Хебба» может усвоить пространственные сигналы и их последовательности. Основная проблема заключалась в том, что автоассоциативная память не в состоянии справиться с отклонениями. Согласно теории, предложенной в нашей книге, кора головного мозга нашла выход из такой ситуации, частично путем складывания автоассоциативных запоминаний в иерархию, а частично – путем использования сложной столбчатой архитектуры. Эта глава почти полностью посвящена иерархической структуре мозга и ее работе, поскольку именно иерархия обусловливает высокую мощность коры головного мозга. Вместо того чтобы пускаться в пространные объяснения, как та или иная клетка учится тем или иным вещам, я хотел бы раскрыть несколько общих принципов обучения в иерархии. У новорожденного кора головного мозга «не знает» фактически ничего. Она не знает его языка, его культуры, его дома, его города, песен, людей, вместе с которыми он будет расти, – ничего. Всю эту информацию – структуру внешнего мира – нужно выучить. Основными составляющими обучения являются формирование классификаций и создание последовательностей. Эти два взаимодополняющих компонента взаимодействуют друг с другом. По мере того как одна зона коры головного мозга учит последовательности, входящие сигналы, которые она посылает в клетки слоя 4 зоной выше в иерархии, меняются. Клетки слоя 4 учатся формировать новые классификации, что изменяет сигнал, проецируемый назад, в слой 1, зоной ниже, а это влияет на последовательности. Основой формирования последовательностей является группирование паттернов, которые относятся к одному и тому же объекту. Один из способов – группировать сигналы, следующие друг за другом. Когда ребенок держит в руке игрушку и медленно ее переворачивает, его мозг может с уверенностью считать, что непрерывно изменяющееся изображение на сетчатке все равно принадлежит одному и тому же объекту, а значит, изменяющийся набор сигналов можно сгруппировать вместе. Иногда нужна внешняя подсказка, чтобы определить, что сигналы принадлежат одному и тому же целому. Чтобы выучить, что яблоки и бананы принадлежат к фруктам, а морковь и картофель – нет, нужен учитель, который помог бы сгруппировать указанные объекты как фрукты. В любом случае ваш мозг создает последовательности сигналов, принадлежащих одному целому. Однако когда зона мозга уже создала последовательности, то входной сигнал, поступающий в следующую область, изменяется. Если раньше он представлял отдельные сигналы, то теперь представляет группу сигналов. Входной сигнал превращается из ноты в мелодию, из букв в слово, из носов в лица и так далее. Поскольку сигналы снизу вверх становятся более объектно-ориентированными, высшая зона коры головного мозга теперь может запоминать последовательности этих объектов высшего порядка. Там, где раньше строились последовательности букв, теперь строятся последовательности слов. Неожиданным результатом процесса обучения является то, что на протяжении повторного обучения представления объектов передвигаются вниз по иерархии. В первые годы вашей жизни «записи» о мире формируются в высших зонах коры головного мозга. Однако по мере вашего дальнейшего обучения они перемещаются во все более низкие зоны иерархии коры головного мозга. Не то чтобы мозг их передвигал туда, просто он должен учить их заново, снова и снова. (Я не хочу сказать, что все запоминания обязательно начинаются с высших зон иерархии коры головного мозга. В реальной жизни усвоение информации выглядит сложнее. Я считаю, что классификация сигналов слоя 4 начинается снизу и передвигается вверх. По мере передвижения формируются последовательности, которые опускаются вниз. Запоминания последовательностей опускаются все ниже и ниже в коре головного мозга.) Когда простые репрезентации передвигаются вниз, высшие зоны коры головного мозга получают возможность учить новые, более сложные, сигналы. Создание и передвижение вниз иерархически организованных воспоминаний можно наблюдать на примере обучения ребенка. Как мы постигаем чтение? Первое, чему нас учат, – распознавать отдельные печатные буквы. Это медленный и сложный процесс, требующий сознательных усилий. Потом мы переходим к распознаванию простых слов. И снова вначале это дается нам с большим трудом и очень медленно, даже если слова состоят из трех-четырех букв. Ребенок может прочесть каждую букву последовательности и произнести каждую из них поочередно, но понадобится достаточно большое количество времени, прежде чем слово будет распознано как слово. Выучив простые слова, мы переходим к изучению многосложных. Сначала мы произносим каждый слог, сцепляя их, как делали это в свое время с простыми словами. После многолетней практики человек способен читать достаточно быстро. Мы достигаем уровня, когда видим не отдельные буквы, а распознаем целые слова и даже фразы с первого взгляда. Дело не в том, что мы стали более быстрыми, а в том, что мы распознаем слова и фразы как целостные единицы. Когда мы читаем все слово, видим ли мы буквы этого слова? И да, и нет. Конечно, сетчатка глаза видит буквы, а значит, и клетки зоны V1 тоже. Однако распознавание букв происходит относительно низко в иерархии коры головного мозга, скажем, в зоне V2 или V4. К тому моменту, когда сигнал попадает в IT, отдельные буквы уже не будут представлены сигналом. То, что вначале требовало деятельности всей зоны коры головного мозга – распознавание отдельных букв, – теперь происходит ближе к сенсорному входному сигналу. Когда запоминания простых объектов (букв) перемещаются вниз по иерархии, высшие зоны коры головного мозга получают возможность учить сложные объекты, такие как слова или фразы. Обучение чтению музыки по нотам может служить еще одним примером. Сначала нужно концентрироваться на каждой ноте. С приобретенным опытом вы начинаете распознавать простые нотные последовательности, затем – целые фразы. После достаточно длительной практики все происходит так, как если бы вы не видели нот вообще. Нотный лист служит вам для того, чтобы помнить общую структуру музыкального фрагмента, а детальные последовательности сохранились в памяти на более низком уровне коры головного мозга. Такой тип обучения происходит как в двигательной, так и в сенсорной зонах. Молодой мозг медленнее распознает входные сигналы, медленнее формирует моторные команды, потому что запоминания, используемые в этих задачах, расположены выше в иерархии. Информация должна пройти весь путь вверх и вниз, возможно, неоднократно, для того, чтобы неясности разрешились. Передвижение сигналов вниз и вверх по иерархической структуре коры головного мозга требует времени. В молодом мозге еще не сформировались сложные последовательности в высших областях, поэтому он не может распознать и отреагировать сложными сигналами. Молодой мозг не может понять высшую структуру внешнего мира. По сравнению со взрослым язык ребенка прост, его музыка незамысловата, и его социальные интеракции тоже просты. Если вы изучаете определенный набор объектов снова и снова, то ваша кора головного мозга смещает в памяти последовательности объектов в нижнюю часть иерархии. Это высвобождает верхние зоны иерархии для изучения более сложных и тонких связей. Согласно моей теории, именно таков путь становления эксперта. Например, я работаю в области компьютерного дизайна, и многие удивляются тому, что я, только взглянув на продукт, уже вижу его недостатки, заложенные в архитектуре. Проработав двадцать пять лет в сфере компьютерного дизайна, я выработал собственную модель всевозможных проблем, связанных с мобильными компьютерами. Точно так же умудренный опытом родитель сможет без труда понять, чем расстроен его ребенок, в то время как новоиспеченный отец будет ломать голову над тем, как же решить возникшую проблему. Опытный менеджер может сразу распознать недостатки и преимущества структуры организации, в то время как начинающий просто не понимает пока этих вещей. Они получают один и тот же входной сигнал, но модель новичка не настолько совершенна, как модель опытного менеджера. В этих и тысяче других примеров мы начинаем с изучения простейшей базовой структуры. Со временем наши знания передвигаются вниз по иерархии, и у наших высших зон появляется возможность освоить структуру более высокого порядка. Именно структуры высшего порядка делают нас опытными. Эксперты и гении обладают мозгом, видящим структуру структуры и сигнал сигналов глубже, чем видят остальные. Можно стать экспертом благодаря практике, но, конечно, кроме этого, существует и генетический компонент таланта и гениальности. Гиппокамп – вершина всех вершинТри большие структуры мозга лежат под оболочкой коры головного мозга и связаны с ней. Это базальные ганглии, мозжечок и гиппокамп. Все три структуры возникли раньше, чем кора головного мозга. В очень грубом приближении мы можем утверждать, что базальные ганглии были примитивной двигательной системой, мозжечок изучал точные временные соотношения событий, а гиппокамп сохранял в памяти конкретные события и места. В определенной степени кора головного мозга присвоила себе функции, изначально принадлежавшие им. Например, человек, родившийся без мозжечка, будет страдать от недостатков координации и вынужден будет прилагать более сознательные усилия при передвижении, но в остальном он будет вполне нормальным. Мы знаем, что кора головного мозга отвечает за все сложные последовательности движений и может непосредственно управлять нашими конечностями. Не то чтобы базальные ганглии не имели никакого значения, просто кора головного мозга узурпировала большую часть моторного контроля. Поэтому я описал общее функционирование коры головного мозга независимо от базальных ганглиев и мозжечка. Некоторые ученые не согласятся с таким моим предположением, тем не менее я использовал его в данной книге и в своей работе вообще. Но вот гиппокамп – другого поля ягода. Это одна из наиболее изученных зон мозга, потому что она является обязательной для формирования новых запоминаний. Если вы потеряете обе половины гиппокампа (как и многие другие составляющие нервной системы, он присутствует и в левом, и в правом полушариях головного мозга), вы утратите способность запоминать новую информацию. Без гиппокампа вы по-прежнему можете разговаривать, ходить, видеть, слышать и даже казаться нормальным в течение небольшого временного промежутка. На самом деле вам причинен глубокий ущерб, ведь вы не можете запомнить ничего нового. Вы будете помнить друга, которого знали до потери гиппокампа, но не сможете запомнить нового человека. Даже если бы вы стали встречаться с доктором пять раз в неделю на протяжении года, все равно каждый последующий раз для вас был бы как первый. У вас бы не было воспоминаний о событиях, которые произошли после того, как вы утратили гиппокамп. Долгие годы у меня не было желания заниматься гиппокампом, поскольку для меня в этом было мало смысла. Гиппокамп является важным для процесса обучения, но не является окончательным хранилищем наших знаний. А вот кора головного мозга таковым является. Классический подход к гиппокампу следующий: здесь формируется память о новых событиях, а затем, через дни, недели, месяцы, эта новая информация перемещается в кору головного мозга. Для меня это звучало как бессмыслица. Мы знаем, что зрительные, слуховые и осязательные сигналы, т. е. сенсорный поток данных, поступают прямо в сенсорные зоны коры головного мозга без предварительного прохождения через гиппокамп. Мне казалось, что сенсорная информация автоматически формирует новые запоминания в коре головного мозга. Зачем нужен гиппокамп в процессе обучения? Как может отдельная структура под видом гиппокампа воспрепятствовать обучению или только позднее передать информацию назад, коре головного мозга? Я решил отложить в сторону гиппокамп, полагая, что однажды его роль прояснится. Так и случилось. Это произошло в конце 2002 года, как раз когда я начал писать эту книгу. Брюно Ольшозен, один из моих коллег из Редвудского института нейрологии, сказал, что связи между гиппокампом и корой головного мозга наводят на мысль о том, что гиппокамп является частью коры головного мозга, а не отдельной структурой. В таком случае гиппокамп расположен на вершине иерархической пирамиды коры головного мозга, это ее самый верхний отдел (см. рис. 6.5). Кора головного мозга появилась на эволюционной сцене, втиснувшись между гиппокампом и остальной частью мозга. Конечно, идея о том, что гиппокамп венчает пирамиду неокортекса, была не нова, но я о ней не знал. Я пообщался с несколькими специалистами в области исследования гиппокампа и попросил их объяснить, как эта структура передает сохраненную информацию в кору головного мозга. Никто мне объяснить не смог. Более того, никто из них не упомянул, что гиппокамп является вершиной пирамиды коры головного мозга, возможно, потому, что гиппокамп не только расположен на вершине корковой пирамиды, но еще и непосредственно связан со многими более старыми частями мозга. Несмотря на это, я сразу понял, что нашел ключ к решению волновавшей меня задачи. Вспомните об информационном потоке, поступающем от ваших глаз, ушей, кожи в кору головного мозга. Каждая зона коры головного мозга пытается понять, что означает полученная информация. Каждая зона пытается объяснить входящий сигнал в рамках известных ей последовательностей. Если она понимает входящий сигнал, то говорит: «Мне это понятно. Это часть объекта, который я уже вижу. Подробности передавать дальше не будем». Если зона не понимает его, она передает его без изменений в более высокую зону иерархии коры головного мозга. Так продолжается до тех пор, пока следующая зона не распознает входной сигнал. Если сигнал является абсолютно новым, он будет подниматься все выше и выше по иерархии. Каждая последующая зона скажет: «Я этого не знаю. Я такого не ожидала. Почему бы вам, высшим областям, не взглянуть на это?» Конечным результатом будет то, что в верхней точке пирамиды коры головного мозга появится информация, которую невозможно понять на основе прошлого опыта. Вы сталкиваетесь с совершенно новым сигналом, которого совсем не ожидали. В повседневной жизни мы встречаем очень много самых разнообразных вещей, информация о которых добирается до верхней части пирамиды. Например, заметка в газете, имя человека, которого вы встретили сегодня утром, автомобильная авария, свидетелем которой вы стали по дороге домой. Вот эта необъясненная и неожиданная информация и попадает в гиппокамп и сохраняется там. Но хранится она там не бесконечно. Позже она либо передается в кору головного мозга, либо исчезает. Я заметил за собой, что с годами мне становится все труднее и труднее запоминать новые вещи. Например, мои дети помнят подробности всех театральных постановок, которые они увидели в прошлом году. Я же не могу этим похвастаться. Возможно, потому, что за всю свою жизнь я увидел такое количество спектаклей, что теперь мне редко попадается что-то на самом деле новое. Новые представления соответствуют воспоминаниям о старых представлениях, возможно, поэтому информация просто не попадает в мой гиппокамп. Если так оно и есть, то приходим к выводу, что, чем больше мы знаем, тем меньше запоминаем. В отличие от коры головного мозга структура гиппокампа гетерогенная, он состоит из нескольких специализированных отделов. Гиппокамп быстро запоминает любые полученные сигналы. Это его уникальная роль, с которой он отлично справляется. Положение гиппокампа на вершине корковой иерархии оптимально для запоминания новых сигналов. У него также наилучшее положение для того, чтобы вызывать эти запоминания и передавать их на хранение в иерархию коры головного мозга. Правда, последний процесс – весьма небыстрый. Вы можете моментально запомнить свежее событие в гиппокампе, но, для того чтобы сохранить его в коре головного мозга навсегда, вам нужно повторять полученный опыт снова и снова, в реальности или мысленно. Альтернативный путь вверх по иерархииУ вашей коры головного мозга существует еще один основной способ передачи информации от одной зоны к другой вверх по иерархии. Этот альтернативный путь начинается с клеток слоя 5, проецирующихся в таламус (в его другую область, не в ту, о которой мы упоминали раньше), а затем из таламуса – в следующую корковую зону. Любые две зоны коры головного мозга, непосредственно связанные друг с другом в иерархии, связаны еще и косвенно – через таламус. Во втором случае информация передается только вверх по иерархии, но не вниз. Когда мы движемся вверх в иерархии коры головного мозга, то существует прямой путь – между областями – и путь косвенный – через таламус. У второго пути есть два режима работы, предопределенные клетками таламуса. Первый режим: путь закрывается и не пропускает информацию. Во втором режиме информация в точности передается от области к области. Этот косвенный путь описали двое ученых, Мюррей Шерман (Нью-Йоркский университет, Стоуни Брук) и Рэй Гиллери (Медицинская школа университета Висконсина). Они полагают, что данный путь может оказаться не менее важным, чем прямой (а может, даже еще более важным, что и было предметом данной главы). У меня есть определенные соображения по поводу того, зачем нужен этот второй путь. Прочтите это слово: воображение. Большинство людей распознают его с одного взгляда, с одной фиксации. Посмотрите на букву «а» в середине этого слова. Теперь посмотрите на хвостик буквы «а». Ваши глаза могут быть все время зафиксированы на одном и том же месте, но в одном случае вы видите слово, в другом – букву, а в третьем – ее хвостик. Смотрите, не отрываясь, на букву и постарайтесь чередовать ваше восприятие слова, буквы и хвостика. Во всех случаях зона V1 получит одну и ту же входящую информацию, но, когда она доберется до высших зон, таких как IT, например, вы будете воспринимать разные вещи, разные уровни детализации. Зона IT знает, как распознать все три объекта. Она может распознать хвостик буквы сам по себе, букву саму по себе и слово целиком с одного взгляда. Когда вы воспринимаете слово целиком, зоны V4, V2, V1 берут на себя детали объекта, а зона IT «узнает» только слово. Во время чтения мы, как правило, не воспринимаем отдельные буквы. Мы воспринимаем слова и фразы. Но если вам захочется, то вы можете распознавать буквы. Мы смещаем акцент внимания очень часто, хотя практически никогда не отдаем себе в этом отчета. Я могу слышать музыку как фон, почти не осознавая этого, но если я постараюсь, то смогу вычленить мелодию ударных инструментов или бас-гитары. В мой мозг поступает один и тот же звук, но я могу сфокусировать свое восприятие. Всякий раз при расчесывании волос возникает сильный внутренний звук, тем не менее вы обычно не осознаете наличие шума. Сосредоточившись, вы сможете очень хорошо услышать данный звук. Все перечисленное – еще один пример сенсорного входящего сигнала, который, как правило, подлежит обработке в нижних слоях коры головного мозга. Но, если вы приложите сознательное усилие, он может попасть и в высшие зоны. Я предполагаю, что альтернативный путь через таламус является тем механизмом, который мы используем, концентрируясь на подробностях, которых, как правило, не замечаем. Он пропускает группирование последовательностей в слое 2, посылая необработанные данные в более высокую зону иерархии. Как доказали биологи, альтернативный путь возможно активизировать двумя способами. Первый – сигналом из высшей зоны самой коры головного мозга. Именно его вы использовали, когда я просил вас обратить внимание на подробности, опускаемые в обычных обстоятельствах (например, хвостик буквы «а» или звук при расчесывании волос). Второй способ – мощный неожиданный сигнал снизу. Если входной сигнал достаточно силен, то он посылает сигнал возбуждения в более высокую область, которая «включает» указанный путь. Например, если бы я показал вам лицо и спросил, что это было, вы бы ответили: «Лицо». Если бы я вам опять показал лицо, но со странной меткой на носу, то вы все равно сразу бы опознали лицо. Затем ваши низшие уровни зрительного восприятия заметили бы: что-то не так. Выявленная ошибка вызывает активизацию дорожки повышенного внимания. Подробности будут передаваться альтернативным путем, пропуская группирование, которое происходит в нормальных условиях. Ваше внимание остановится на метке. Теперь вы видите не только лицо, но и метку. Если она выглядит достаточно необычно, то может полностью завладеть вашим вниманием. Так необычные события быстро попадают в зону нашего внимания. Именно поэтому нам очень трудно не обращать внимания на физические изъяны или какие-либо необычные сигналы. Наш мозг делает это автоматически. Однако очень часто сообщения об ошибке оказываются недостаточно сильными для того, чтобы открыть альтернативный путь. Поэтому иногда мы не замечаем орфографических ошибок в словах, которые читаем. Заключительные размышленияДля того чтобы найти и разработать новую научную модель, мы должны искать наиболее простые подходы, которые могли бы объединить и объяснить множество разрозненных фактов. Неизбежным следствием такого процесса является то, что по инерции мы можем сильно упрощать ситуацию. Можно пропустить важные детали и ошибочно интерпретировать факты. Но если модель в общем будет принята, то можно будет ее усовершенствовать и исправить ошибки. Постепенно станет ясно, где мы зашли слишком далеко, где мы копали недостаточно глубоко, а где ошибались. В этой главе я изложил несколько гипотез по поводу работы коры головного мозга. Полагаю, некоторые из них окажутся ошибочными, возможно также, что все гипотезы будут пересмотрены. Осталось очень много деталей, которые не были мной даже упомянуты. Мозг сам по себе очень сложен. Нейробиологи, читающие эту книгу, знают, что я представил только схематическую характеристику сложнейшего биологического устройства – живого человеческого мозга. Тем не менее я считаю, что предложенная мной теоретическая модель в целом выглядит убедительно. Я надеюсь, что, несмотря на изменения по мере появления новых данных, мои основные теоретические положения все-таки окажутся верными. И наконец, возможно, вы все еще не можете поверить в то, что простая, но обширная система запоминания в самом деле может отвечать за всю деятельность человека. Разве возможно, чтобы вы и я были просто иерархической системой запоминания? Разве возможно, чтобы наши жизни, верования и амбиции хранились в триллионах крошечных синапсов? В 1984 году я начал работать в сфере создания компьютерных программ. Конечно, я писал небольшие программы и раньше, но это был мой первый опыт программирования компьютера с графическим интерфейсом для пользователя. Это был первый опыт работы над большими и сложными приложениями. Я создавал программное обеспечение для операционной системы, созданной Grid Systems. Там были окна, много шрифтов, пунктов меню. Операционная система указанной компании на то время была очень продвинутой. Однажды я был поражен мыслью, что занимаюсь чем-то практически невозможным. Как программист, я мог писать только одну строку кода за другой. Потом я группировал строки кодов в блоки, называемые подпрограммами. Затем подпрограммы объединялись в модули. Объединение модулей давало в результате приложение. Программа электронной таблицы, над которой я работал, состояла из такого большого количества подпрограмм и модулей, что ни один человек не был в состоянии понять ее всю. Она была очень сложной. Тем не менее одна строка кода в ней мало что делает. Требовалось несколько строк кода, чтобы отобразить только один пиксель на экране. Чтобы заполнить весь экран для таблицы, компьютеру надо было выполнить миллионы команд, переданных сотнями подпрограмм в повторяющемся и рекурсивном порядке. Все это было настолько сложно, что было невозможно предвидеть, что происходит, когда программа работает. Я был поражен: насколько нереалистичным казалось то, что при работе программы экран будет заполнен чуть ли не мгновенно. Внешне плоды работы программы выглядели как таблицы с числами, ярлыки, текст, графика. Она работала как электронная таблица. Однако я знал, что происходит внутри компьютера, процессор которого постоянно выполняет одну и ту же простую операцию. Трудно было поверить в то, что компьютер может сориентироваться в лабиринте модулей и подпрограмм и выполнить команды настолько быстро. Если бы у меня не было предварительных знаний, я бы попросту не поверил, что все это будет работать. Если бы кто-нибудь изобрел компьютер с графическим интерфейсом для пользователя и приложением электронных таблиц и принес бы мне свое изобретение на бумаге, я бы отклонил его как не имеющее практического применения. Я бы возразил: такой машине понадобится вечность, чтобы что-либо сделать. И тут мне стало стыдно за себя, ведь на самом деле она работает. Я понял, что мое интуитивное представление о скорости процессора и мое интуитивное представление о мощности иерархического строения не соответствуют действительности. Здесь уместно привести аналогию с корой головного мозга. Мозг не состоит из сверхбыстрых компонентов, правила, которых он придерживается в своей работе не так уж сложны. Однако есть одно «но»: кора головного мозга обладает иерархической структурой, она состоит из миллиардов нейронов и триллионов синапсов. Нам трудно представить, как такая логически простая, но по численности обширная система памяти может создавать наше сознание, язык, культуру, искусство, книги, науку и технологии. Однако это происходит, потому что наше интуитивное представление о способностях коры головного мозга и мощностях ее иерархической структуры является неадекватным. Кора головного мозга на самом деле работает. В этом нет никакой магии. Это доступно нашему пониманию. И точно так же, как в случае с компьютером, в конечном счете мы сможем создать разумные машины, работающие по тому же принципу. Примечания:1 В Силиконовой долине, которая протянулась на 40 км от г. Сан-Хосе на юго-западе до г. Пало-Алто на северо-западе США, расположены ведущие американские высокотехнологичные фирмы. – Примеч. ред. 15 Фотообъектив типа «рыбий глаз» имеет поле зрения, близкое к 180 градусам, внося искажения, которые можно контролировать только пользуясь зеркальным видоискателем. – Примеч. ред. 16 Фотообъектив типа «рыбий глаз» имеет поле зрения, близкое к 180 градусам, внося искажения, которые можно контролировать только пользуясь зеркальным видоискателем. – Примеч. ред. |
|
|||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Наверх |
||||
|
|
||||